Theo Rein, München
FKBP5 bei Depression
Moderne Arzneimittelentwicklung leitet ihre Zielstrukturen aus einem verbesserten Verständnis der molekularen Vorgänge der zu behandelnden Krankheit ab. Die hohe Komplexität psychiatrischer Krankheiten wie der Depression stellt eine besondere Herausforderung für die Entschlüsselung dieser grundlegenden Vorgänge dar. Es ist deswegen nicht überraschend, dass die enormen Anstrengungen zur Aufklärung der molekularen Prozesse der Depression auf verschiedenen Ebenen, etwa des Genoms, Epigenoms, Transkriptoms, Proteoms, Metaboloms etc., noch kein einheitliches Bild ergeben haben.
Stress, Depression und die Glucocorticoid-Rezeptor-Hypothese
Auch wenn allgemein akzeptiert ist, dass Dysfunktionen unterschiedlicher molekularer Signalwege zur Entstehung der Depression beitragen, so gilt doch die Rolle von Stress als besonders bedeutsam. Die molekularen Grundlagen zur Verarbeitung von Stress sind wiederum multifaktoriell. Eine zentrale Stellung nimmt hierbei der Glucocorticoid-Rezeptor (GR) ein. Er ist Teil der Stresshormonachse und vermittelt die zellulären Effekte des Stresshormons Cortisol. Die klinische Beobachtung des Einflusses von Stress bei der Depression und der Korrelation der Cortisolspiegel mit dem Behandlungserfolg führten zur Glucocorticoid-Rezeptor-Hypothese der Depression [24]. Dieser Hypothese zufolge ist eine Fehlfunktion des GR entscheidend für die Entstehung dieser Krankheit.
FKBP51 ist ein Stressregulator
Die Glucocorticoid-Rezeptor-Hypothese lenkte das Augenmerk der molekularen Psychiatrie auf die Suche nach Faktoren, die für die korrekte Funktion des GR verantwortlich zeichnen. In Abwesenheit von Cortisol befindet sich der GR in einem zytosolischen Heterokomplex aus verschiedenen Chaperon- und Cochaperon-Proteinen. Die Zusammensetzung dieser Chaperon-Proteine wiederum bestimmt die Bindungsaffinität des Rezeptors zu Cortisol und damit seine Fähigkeit, auf die Erhöhung des Stresshormons zu reagieren [21]. Unter den Chaperonen und Cochaperonen des GR-Heterokomplexes weckte das FK506-bindende Protein 51 (FKBP51) besonderes Interesse. Dieses Produkt des FKBP5-Gens wurde bei Totenkopfäffchen, einem Primatenmodell der Glucocorticoid-Resistenz [7], in erhöhter Konzentration gefunden [9]. Die Ergebnisse weiterer Studien konnten zeigen, dass FKBP51 ein starker Inhibitor des GR ist und dadurch auch seine eigene Synthese hemmt [9, 35, 39, 44]. Es führt unter anderem zu einer geringeren Bindungsaffinität des GR zu Cortisol [44], die aus dem Dexamethason-Suppressionstest auch bei depressiven Patienten für den GR abgeleitet worden war [32].
Vom Kandidatengen zum Kandidaten für die Arzneimittelentwicklung
Auf der Basis der Etablierung von FKBP51 als GR-Inhibitor fand dessen Gen FKBP5 Eingang in eine Kandidatengen-Assoziationsstudie, die eine Korrelation bestimmter Varianten dieses Gens mit erhöhtem Rezidivrisiko für eine Depression einerseits und einem schnelleren Ansprechen auf Antidepressiva andererseits entdeckte. Diese Genvarianten waren außerdem mit veränderten Proteinspiegeln von FKBP51 assoziiert [6]. Diese Korrelationen wurden in einigen, aber nicht allen nachfolgenden genetischen Studien repliziert, insbesondere nicht in den genomweiten [12, 25, 45]. Auch wenn methodische Gründe nicht vollkommen ausgeschlossen werden können, spiegelt sich darin wahrscheinlich die molekulare Komplexität der Depression wider. Die Bedeutung von FKBP5 für die Regulation der Stresshormonachse, Depressions-ähnliches Verhalten und die Wirkung von Antidepressiva wurde in mehreren tierexperimentellen Studien belegt [12]. Die Gesamtheit dieser klinischen und präklinischen Arbeiten begründete die Erwartung, dass FKBP51 ein vielversprechendes Zielprotein für die Entwicklung neuartiger Antidepressiva sein könnte [38].
FKBP51 reguliert nicht nur Stress
Nach der Entdeckung von FKBP51 als Stressregulator und der Assoziation seines Gens FKBP5 mit Wiederauftreten und Behandlung von Depression wurde die Regulation und Funktion des Gens und des Proteins intensiv erforscht. Dabei wurden vielfältige Proteininteraktionen und von FKBP51 kontrollierte Signalwege entdeckt, ebenso wie zahlreiche genetische Assoziationen und Verbindungen zu physiologischen und pathophysiologischen Prozessen [22, 35, 41, 49].
FKBP51 in diversen Signalwegen
FKBP51 findet sich auch in Komplexen mit den anderen Steroidrezeptoren und bestimmt insbesondere die Aktivität des Mineralocorticoid-Rezeptors und des Progesteron-Rezeptors [4, 39]. Außerdem bildet FKBP51 regulatorische Komplexe mit Proteinen aus dem Bereich des Entzündungs- und Immungeschehens aus. Dazu gehören nicht nur der GR als bekannter Immunmodulator, sondern beispielsweise auch Proteine des NFκB-Signalwegs, Calcineurin- und TRAF(TNF-Rezeptor-assoziierte Faktor)-Proteine [12, 22, 49]. FKBP51 gestaltet weiterhin die Ausbildung eines regulatorischen Komplexes mit der Proteinkinase B (Akt), der zu deren Dephosphorylierung und damit Inaktivierung führt [10, 33]. In ähnlicher Weise bestimmt FKBP51 über die Organisation anderer Proteinkomplexe den Phosphorylierungsstatus und damit die Aktivität von Proteinen, beispielsweise der DNA-Methyltransferase 1 (DNMT1) [15], Glykogensynthase-Kinase 3β (GSK-3 β) [16], AS160 (Akt substrate of 160 kD) [3], Beclin1 [17] und SKP2 (S-Phase kinase associated protein 2) [18]. FKBP51 ist also ein multifunktionales Gerüstprotein, das über all diese Proteinassoziationen die molekulare Stressantwort mit Signalwegen der Autophagie, Zellproliferation, Metabolismus, Genprogrammierung usw. verbindet, die sehr häufig rückgekoppelt sind wie andernorts ausgeführt [35] (Abb. 1).
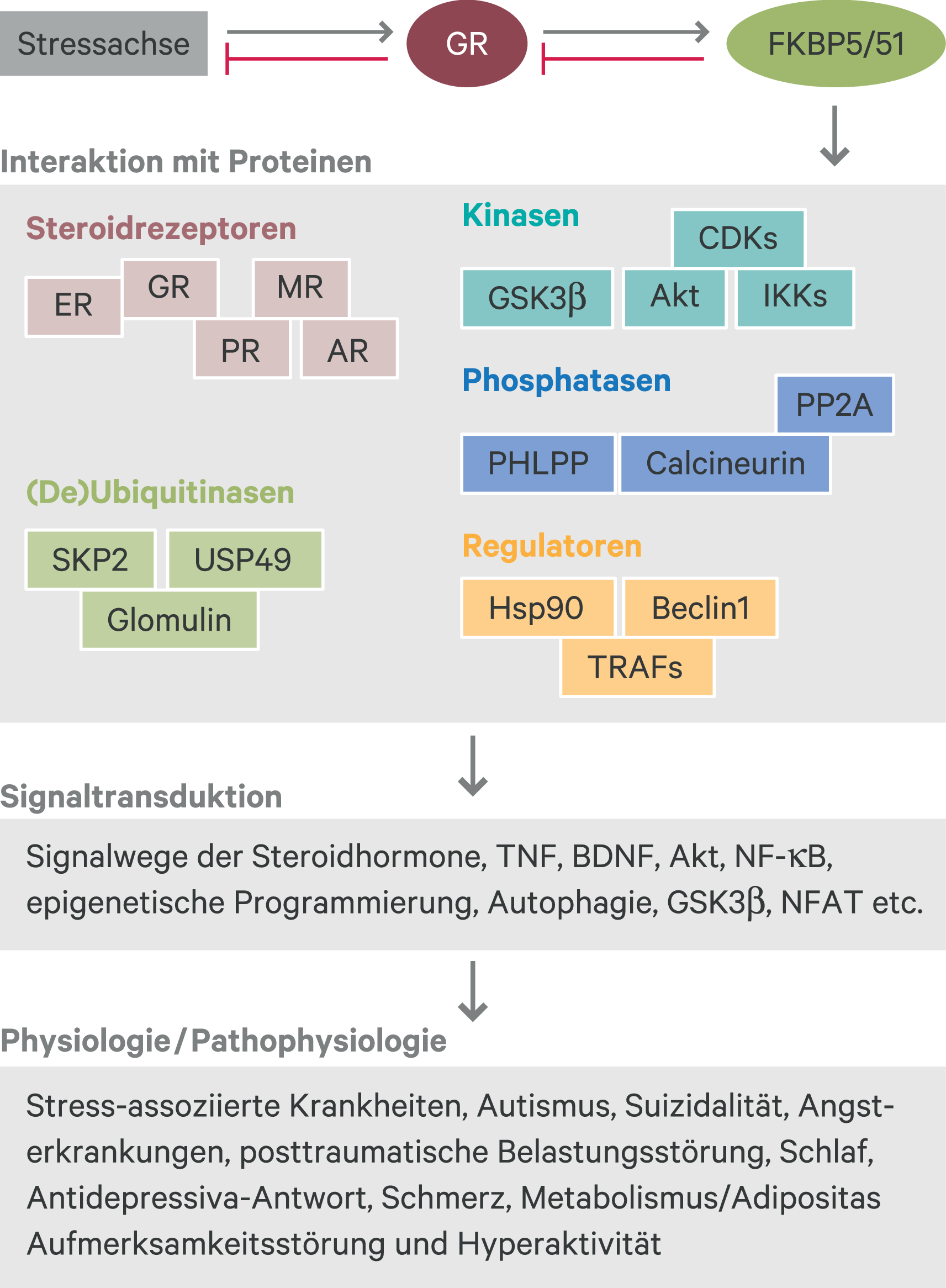
Abb. 1. Verflechtung von FKBP51 mit Stress, diversen Signalwegen und weiteren physiologischen Prozessen Stress aktiviert über Cortisol und den GR das Gen FKBP5, dessen Produkt FKBP51 den GR inhibiert. Die so eingestellten FKBP51-Spiegel sind über Proteinwechselwirkungen mit zahlreichen Signalwegen verknüpft, deren Aktivität wiederum verschiedenste Aspekte der Physiologie und Pathophysiologie bestimmt. Die Abbildung gibt jeweils nur eine Auswahl wieder. Akt: Proteinkinase B; AR: Androgen-Rezeptor; BDNF: Brain-derived neurotrophic factor; CDKs: Cyclin-dependent kinases; ER: Estrogen-Rezeptor; GR: Glucocorticoid-Rezeptor; GSK3β: Glykogensynthase-Kinase 3β; Hsp90: Heat shock protein 90; IKKs: IκB kinase; MR: Mineralocorticoid-Rezeptor; NFAT: Nuclear factor of activated T-cells; NF-κB: Nuclear factor kappa-light-chain-enhancer of activated B cells; PHLPP: Pleckstrin homology domain leucine-rich repeat protein phosphatase; PP2A: Protein phosphatase 2; PR: Progesteron-Rezeptor; SKP2: S-phase kinase associated protein 2; TNF: Tumor necrosis factor; TRAFs: Tumor necrosis factor receptor associated factors; USP49: Ubiquitin specific peptidase 49
Physiologische Rolle von FKBP51
Zahlreiche Arbeiten von zellulären und tierexperimentellen Ansätzen bis hin zu humangenetischen und klinischen Studien zeigen eine Funktion von FKBP51 in vielen physiologischen und pathophysiologischen Zusammenhängen. Dazu gehören neben der Stressregulation und Depression auch Sucht, Autismus, Schizophrenie, Chorea Huntington, Suizidalität, Angsterkrankungen, posttraumatische Belastungsstörung (PTSD), Schlafstörungen, die Förderung der Antidepressiva-Antwort, die Interaktion mit Zytostatika, chronischer Schmerz, Adipositas sowie Aufmerksamkeitsstörung und Hyperaktivität [2, 8, 12, 22] (Abb. 1). Es ist höchst unwahrscheinlich, dass sich die physiologischen Effekte jeweils durch nur eine bestimmte Proteininteraktion von FKBP51 erklären lassen. Die Verknüpfung mit dem GR liegt bei der Stressregulation und Stress-assoziierten Krankheiten nahe, aber beispielsweise die epigenetische Programmierung der zugrunde liegenden Gene wie etwa des Brain-derived neurotrophic factors (BDNF) könnte durch die Wechselwirkung von FKBP51 mit der DNMT1 mitbestimmt sein. Die Effekte von Antidepressiva konnten in Zusammenhang mit den FKBP51-regulierten Signalwegen gebracht werden: Es zeigte sich, dass FKBP51 die Effekte von konventionellen Antidepressiva wie Paroxetin, Fluoxetin und Amitriptylin auf Autophagie, GSK3β und DNMT1-BDNF beförderte [15, 16, 17]. Kürzlich wurde außerdem gezeigt, dass das als schnell wirkendes Antidepressivum diskutierte Ketamin für die Stimulierung der Ausschüttung von BDNF ebenfalls die Anwesenheit von FKBP51 benötigt [1]. In einer Genexpressionsanalyse im Striatum und Hippocampus der Maus wurde eine erhöhte FKBP5-Transkription zwei bis vier Stunden nach der Applizierung von Ketamin entdeckt [11]. Die Gabe von Ketamin erhöhte außerdem die mRNA-Spiegel von FKBP5 in peripheren mononukleären Blutzellen von Depressionspatienten [26].
Entwicklung von FKBP51-Liganden
Typischerweise zielt die Entwicklung eines neuen Arzneimittels auf die Inhibierung der biochemischen Funktion des entsprechenden Zielproteins ab, zum Beispiel die spezifische Blockade einer bestimmten Adenosintriphosphatase (ATPase). Dabei werden in der Regel Substanzen entwickelt, die in die Bindungstasche für die Reaktanten der katalysierten biochemischen Umsetzung im Zielprotein eingepasst werden, beispielsweise die ATP-Bindungstasche. FKBP51 hat eine modulare Struktur, die unter anderen die Domäne für die Hsp(Heat shock protein)90-Bindung und die Domäne für die Katalyse der cis-trans-Isomerisierung von Prolylbindungen in Proteinen (FK1-Domäne) umfasst [41]. Während die Interaktion mit Hsp90 notwendig für die Wirkung von FKBP51 auf den GR ist, legen die Ergebnisse von Mutationsstudien nahe, dass die Funktion der Peptidylprolylisomerase (PPIase) hierfür verzichtbar ist [44]. Dennoch konnte die flache Bindungsstelle der PPIase aus verschiedenen Gründen als geeignete Zielstruktur für wirksame FKBP51-Liganden angesehen werden: Mit Tacrolimus (FK506) und Sirolimus (Rapamycin) gibt es zwei klinisch validierte Substanzen, die mit hoher Affinität in die PPIase-Tasche binden [47]. Diese Strukturen sind als Ausgangspunkt für medizinalchemische Anpassungen und Weiterentwicklungen geeignet [38]. Die PPIase-Aktivität kann in Hochdurchsatzverfahren gemessen werden, sodass die Affinität der Ligandenbindung präzise und effizient bestimmt werden kann. Darüber hinaus ist zwar die PPIase-Aktivität von FKBP51 für die Inhibierung des GR verzichtbar, nicht aber die PPIase-Domäne. Es ist deshalb sehr wahrscheinlich, dass Liganden entwickelt werden können, die die Wechselwirkung dieser Domäne mit anderen Proteinen wie dem GR stören. Das ursprüngliche Konzept verfolgte also das Ziel, durch die Inhibition der Funktion des GR-Inhibitors FKBP51 die Funktion des GR wiederherzustellen (Abb. 2).
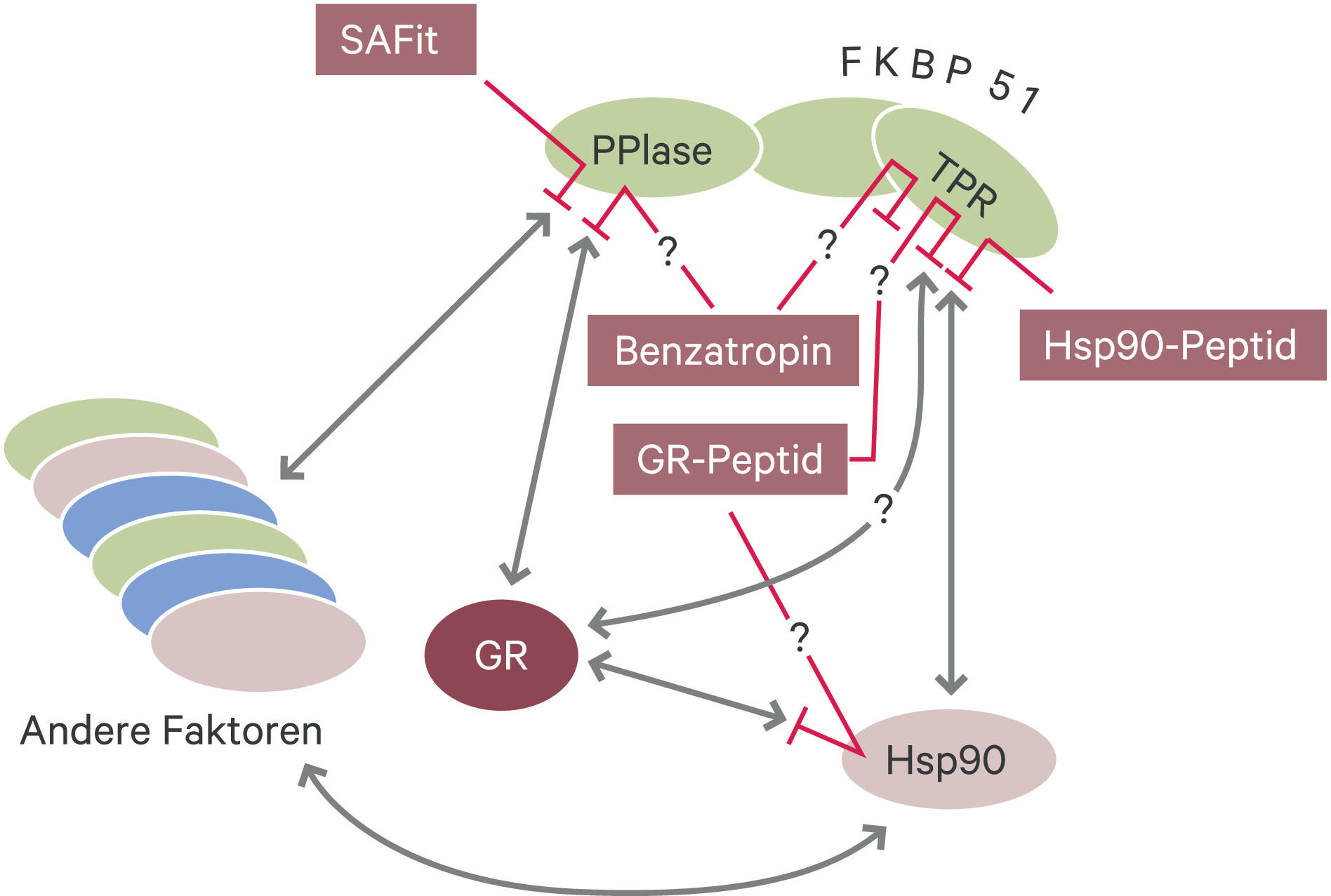
Abb. 2. Strategien zur Inhibition von FKBP51 Von FKBP51 sind zwei für die zahlreichen Wechselwirkungen mit anderen Proteinen entscheidende Domänen bekannt, die PPIase-Domäne und die TPR(Tetratricopeptide repeat)-Domäne. Letztere ist für die Bindung an das zentrale Chaperon Hsp90 verantwortlich und ermöglicht darüber die Einbindung in weitere Proteinkomplexe. (1) Inhibitoren der PPIase-Aktivität von FKBP51 wie SAFit1 und SAFit2 üben ihren Effekt höchstwahrscheinlich über die Störung der Interaktion mit anderen Proteinen aus. Inwieweit sich über diese Strategie Inhibitoren entwickeln lassen, die spezifisch nur einen Teil der FKBP51-Interaktionen blockieren, muss noch gezeigt werden. (2) Benzatropin bindet an FKBP51 ohne die PPIase-Aktivität zu hemmen und inhibiert den Einbau in den Komplex aus Hsp90 und GR über einen noch aufzuklärenden Mechanismus. (3) Von Hsp90 oder GR abgeleitete Peptide repräsentieren weitere Strategien zur gezielten Hemmung des FKBP51-Effekts auf den GR. Das vorgeschlagene Hsp90-Peptid blockiert die TPR-Bindestelle, sodass unter anderem die Frage der Selektivität unter den verschiedenen TPR-Proteinen gelöst werden muss. Die Bindestelle des wirksamen GR-Peptids muss noch kartiert werden.
SAFit1 und SAFit2
Die Entwicklung von FKBP51-Liganden sah sich der besonderen Herausforderung gegenüber, eine Spezifität gegenüber dem zu FKBP51 hoch homologen FKBP52 zu erreichen. FKBP52 tritt regelmäßig als funktionaler Antagonist von FKBP51 auf, zum Beispiel als Stimulator der GR-Aktivität [44, 49]. Die zunächst synthetisierten Substanzen nahmen einen für FKBP12 entwickelten Liganden als Ausgangspunkt, wodurch noch keine ausreichend hohen Affinitäten für FKBP51 erreicht werden konnten und auch keine Selektivität gegenüber FKBP52 [19, 20]. Der Schlüssel für die Entwicklung selektiver Liganden liegt in der unterschiedlichen konformationellen Dynamik von FKBP51 und FKBP52 [13, 27]. Während in den Kristallstrukturen der beiden hoch homologen Proteine die PPIase-Bindungstaschen kaum unterscheidbar sind [40, 41, 46], zeigt sich durch NMR(Nuclear magnetic resonance)-spektroskopische und andere Analysen ein weiterer und seltener konformationeller Zustand für FKBP51, der von SAFit (Selective antagonists of FKBP51 by induced fit) gebunden und stabilisiert wird [13, 27, 30, 43]. Im ungebundenen Zustand stehen die konformationellen Zustände im dynamischen Gleichgewicht [27, 30]. Die Entdeckung dieser Konformation ist die Grundlage für die gezielte Entwicklung weiterer Liganden mit geringerem Molekulargewicht und verbesserten physikochemischen Eigenschaften [5, 14]. SAFit1 und SAFit2 wurden bereits erfolgreich in mehreren tierexperimentellen Arbeiten eingesetzt und haben als analytisches Werkzeug wesentlich zur Aufklärung der zuvor beschriebenen Funktionen von FKBP51 beigetragen [5, 22] (Tab. 1).
Tab. 1. Experimentelle Hinweise auf antidepressives Potenzial von Substanzen, die an FKBP51 binden oder dessen Funktion stören
|
Substanz |
Experimenteller Hinweis |
Referenz |
|
Benzatropin |
Verhinderte in Zellkulturen den Einfluss von FKBP51 auf die Kerntranslokation und transkriptionelle Aktivität des GR. Die Bindestelle an FKBP51 ist noch nicht kartiert. |
[36] |
|
GR-Peptid |
Verhinderte die Bindung von FKBP51 an GR; erhöhte Phosphorylierung und Kerntranslokation des GR. Injektion in die Amygdala reduzierte Angstverhalten im Tierexperiment (Maus). |
[31] |
|
SAFit2 |
Intraperitoneale Injektion bewirkte im erzwungenen Schwimmtest bei Mäusen Verhaltensänderungen wie klassische Antidepressiva. |
[13] |
|
SAFit2 |
Mikroinjektion in basolaterale Amygdala reduzierte angstähnliches Verhalten bei Mäusen, ebenso wie die periphere Injektion. |
[23] |
|
SAFit2 |
Kobehandlung von Mäusen verringerte die Effekte des Antidepressivums Escitalopram in angstähnlichem Verhalten, verstärkte aber die Effekte im erzwungenen Schwimmtest. |
[34] |
SAFit: Selective antagonists of FKBP51 by induced fit (niedermolekulare Liganden für FKBP51)
Weitere Strategien zur Inhibierung von FKBP51
SAFit1 und SAFit2 sind zweifelsohne die mit Abstand am weitesten entwickelten und verwendeten Liganden für FKBP51 [28]. Auf der Suche nach alternativen Strategien zur Modulierung von FKBP51 wurde bei der Analyse von 1280 pharmakologisch aktiven Substanzen Benzatropin als FKBP51-Inhibitor entdeckt [36]. Benzatropin bindet an einer noch zu kartierenden Stelle an FKBP51 und blockiert dessen Effekt auf den GR dadurch, dass es die Assoziation mit dem Komplex aus GR und Hsp90 verhindert. Es blockiert hingegen nicht die PPIase-Aktivität von FKBP51 und zeigt somit ein von SAFit verschiedenes Wirkprinzip (Abb. 2, Tab. 1). Benzatropin ist als anticholinerges Arzneimittel unter dem Handelsnamen Cogentin® bekannt und wird unter anderem bei Morbus Parkinson eingesetzt. Weil es somit auch die Blut-Hirn-Schranke überwinden kann, wurde es als Ausgangspunkt für eine medizinalchemische Optimierung für die Steuerung der FKBP51-Aktivität vorgeschlagen [36].
Inhibierung der Wechselwirkung mit dem Chaperon Hsp90
Neben der PPIase-Domäne ist auch die Domäne für die Wechselwirkung mit dem zentralen Chaperon Hsp90 für den Effekt von FKBP51 auf den GR essenziell [44]. Entsprechend ihrer Struktur wird diese Domäne auch als TPR(Tetratricopeptide repeat)-Domäne bezeichnet, die sich in mehreren Proteinen findet [48]. Theoretisch könnte die Funktion von FKBP51 im Hinblick auf den GR also auch durch Blockierung seiner TPR-Domäne gestört werden. Dieses Prinzip war bereits vor mehr als 20 Jahren als Möglichkeit zur pharmakologischen Beeinflussung von TPR-Proteinen wie FKBP51 vorgeschlagen worden [37]. Kürzlich wurde diese Idee durch die eingehende Charakterisierung der Bindungsmodalitäten eines von Hsp90 abgeleiteten Peptids an die TPR-Domäne von FKBP51 wieder aufgegriffen [29] (Abb. 2). Wie bei anderen TPR-Domänen leisten ionische Wechselwirkungen einen wichtigen Beitrag zu dieser Bindung. Zu erwartende Schwierigkeiten bei der weiteren Verfolgung dieser Idee sind also neben der Unterscheidung der verschiedenen TPR-Proteine auch die konkurrierende Bindung an andere positiv geladene Strukturen in der Zelle sowie die Überwindung der Zellmembran und der Blut-Hirn-Schranke.
Inhibierung der Wechselwirkung mit dem GR
Eine weitere Strategie zur gezielten Beeinflussung der Aktivität von FKBP51 auf den GR erwuchs aus der Charakterisierung seiner Kontaktstelle am GR [31]. Aus dieser Kontaktstelle wurde eine kurze Peptidsequenz abgeleitet, die anstelle des GR an FKBP51 binden soll, um den Effekt auf den GR zu blockieren [31]. Dieses Peptid zeigte positive Effekte in einem PTSD-Modell in der Maus, und verbesserte die molekulare GR-Funktion auf verschiedenen Ebenen, unter anderem durch eine Erhöhung der GR-Bindung an FKBP52 [31]. Ob dieser interessante Mechanismus wie vorgeschlagen pharmakologisch nutzbar sein wird, müssen künftige Studien zeigen. In diesen muss beispielsweise geklärt werden, ob die beschriebene GR-Peptidsequenz tatsächlich direkt an FKBP51 (in der TPR-Domäne) bindet, oder nicht doch über Hsp90 (Abb. 2, Tab. 1). Um das Peptid über die Zellmembran zu bringen, wurde es in einer präklinischen Studie mit der Transduktionsdomäne des HIV1-TAT(Tat protein of human immunodeficiency virus type 1)-Proteins fusioniert. Derzeit ist noch offen, inwieweit diese in der Grundlagenforschung erfolgreiche Methode ihren Weg in die klinische Therapie finden kann [42, 50].
Diskussion und Ausblick
Angesichts der vielfältigen Funktionen von FKBP51 wird häufig die Frage gestellt, ob es denn als Zielstruktur für die Arzneimittelentwicklung, zum Beispiel für die Behandlung von Depression, tauge. Glücklicherweise können bisher weder aus der kompletten Ausschaltung von FKBP5 im Mausmodell noch aus der Gabe von Inhibitoren negative Auswirkungen abgeleitet werden. Die Eignung von FKBP51 als Zielstruktur wird auch nicht dadurch infrage gestellt, dass Antidepressiva in FKBP5-depletierten Mäusen ihren Effekt verlieren und erhöhte Spiegel von FKBP51 bei an Depression Erkrankten mit einer verbesserten Antidepressiva-Wirkung einhergehen. Möglicherweise sind FKBP51-Inhibitoren aber nur für eine Untergruppe von depressiven Patienten geeignet. Da FKBP51 als Relais für verschiedene Signalwege fungiert, können über FKBP51 mehrere physiologische Prozesse angesprochen werden. Die wahrscheinlichste Erklärung für die Wirkung PPIase-basierter FKBP51-Liganden ist die Interferenz mit Protein-Wechselwirkungen von FKBP51. Für die Entwicklung gezielter einsetzbarer Liganden ist ein besseres Verständnis darüber notwendig, welche FKBP51-Proteininteraktionen und Signalwegeffekte welchem physiologischen Effekt zugrunde liegen. Dies könnte das Design von PPIase-Liganden erleichtern, die nur einen Teil der FKBP51-Interaktionen stören, und damit das gezielte Ansprechen bestimmter Signalwege ermöglichen (Abb. 2). Daraus ergibt sich das Potenzial für die Entwicklung nicht nur von Antidepressiva, sondern auch von Substanzen für weitere Indikationen. Auf dem Weg dahin könnten die peptidischen FKBP51-Inhibitoren als analytisches Werkzeug von großem Nutzen sein, auch wenn sie selbst für die Entwicklung eines Arzneimittels eher ungeeignet erscheinen.
Interessenkonflikterklärung
Dr. Theo Rein fungiert als Editor für das Journal of Cellular Biochemistry.
Literatur
1. Anderzhanova E, Hafner K, Genewsky AJ, Soliman A, et al. The stress susceptibility factor FKBP51 controls S-ketamine-evoked release of mBDNF in the prefrontal cortex of mice. Neurobiol Stress 2020;13:100239.
2. Bailus BJ, Scheeler SM, Simons J, Sanchez MA, et al. Modulating FKBP5/FKBP51 and autophagy lowers HTT (huntingtin) levels. Autophagy 2021;17:4119–40.
3. Balsevich G, Häusl AS, Meyer CW, Karamihalev S, et al. Stress-responsive FKBP51 regulates AKT2-AS160 signaling and metabolic function. Nat Commun 2017;8:1725.
4. Barent RL, Nair SC, Carr DC, Ruan Y, et al. Analysis of FKBP51/FKBP52 chimeras and mutants for Hsp90 binding and association with progesterone receptor complexes. Mol Endocrinol 1998;12:342–54.
5. Bauder M, Meyners C, Purder PL, Merz S, et al. Structure-based design of high-affinity macrocyclic FKBP51 inhibitors. J Med Chem 2021;64:3320–49.
6. Binder EB, Salyakina D, Lichtner P, Wochnik GM, et al. Polymorphisms in FKBP5 are associated with increased recurrence of depressive episodes and rapid response to antidepressant treatment. Nat Genet 2004;36:1319–25.
7. Chrousos GP, Renquist D, Brandon D, Eil C, et al. Glucocorticoid hormone resistance during primate evolution: receptor-mediated mechanisms. Proc Natl Acad Sci U S A 1982;79:2036–40.
8. Connelly KL, Wolsh CC, Barr JL, Bauder M, et al. Sex differences in the effect of the FKBP5 inhibitor SAFit2 on anxiety and stress-induced reinstatement following cocaine self-administration. Neurobiol Stress 2020;13:100232.
9. Denny WB, Valentine DL, Reynolds PD, Smith DF, et al. Squirrel monkey immunophilin FKBP51 is a potent inhibitor of glucocorticoid receptor binding. Endocrinology 2000;141:4107–13.
10. Fabian AK, Mährz A, Neimanis S, Biondi RM, et al. InterAKTions with FKBPs – mutational and pharmacological exploration. PLoS One 2013;8:e57508.
11. Ficek J, Zygmunt M, Piechota M, Hoinkis D, et al. Molecular profile of dissociative drug ketamine in relation to its rapid antidepressant action. BMC Genomics 2016;17:362.
12. Fries GR, Gassen NC, Rein T. The FKBP51 glucocorticoid receptor co-chaperone: regulation, function, and implications in health and disease. Int J Mol Sci 2017;18:E2614.
13. Gaali S, Kirschner A, Cuboni S, Hartmann J, et al. Selective inhibitors of the FK506-binding protein 51 by induced fit. Nat Chem Biol 2015;11:33–7.
14. Gabani BB, Sulochana SP, Siddesh AHA, Kiran V, et al. Validated LC-MS/MS method for simultaneous quantitation of SAFit-1 and SAFit-2 in mice plasma: application to a pharmacokinetic study. Drug Res (Stuttg) 2020;70:325–32.
15. Gassen NC, Fries GR, Zannas AS, Hartmann J, et al. Chaperoning epigenetics: FKBP51 decreases the activity of DNMT1 and mediates epigenetic effects of the antidepressant paroxetine. Sci Signal 2015;8:ra119.
16. Gassen NC, Hartmann J, Zannas AS, Kretzschmar A, et al. FKBP51 inhibits GSK3beta and augments the effects of distinct psychotropic medications. Mol Psychiatry 2016;21:277–89.
17. Gassen NC, Hartmann J, Zschocke J, Stepan J, et al. Association of FKBP51 with priming of autophagy pathways and mediation of antidepressant treatment response: evidence in cells, mice, and humans. PLoS Med 2014;11:e1001755.
18. Gassen NC, Niemeyer D, Muth D, Corman VM, et al. SKP2 attenuates autophagy through Beclin1-ubiquitination and its inhibition reduces MERS-Coronavirus infection. Nat Commun 2019;10:5770.
19. Gopalakrishnan R, Kozany C, Gaali S, Kress C, et al. Evaluation of synthetic FK506 analogues as ligands for the FK506-binding proteins 51 and 52. J Med Chem 2012;55:4114–22.
20. Gopalakrishnan R, Kozany C, Wang Y, Schneider S, et al. Exploration of pipecolate sulfonamides as binders of the FK506-binding proteins 51 and 52. J Med Chem 2012;55:4123–31.
21. Grad I, Picard D. The glucocorticoid responses are shaped by molecular chaperones. Mol Cell Endocrinol 2007;275:2–12.
22. Hähle A, Merz S, Meyners C, Hausch F. The many faces of FKBP51. Biomolecules 2019;9:E35.
23. Hartmann J, Wagner KV, Gaali S, Kirschner A, et al. Pharmacological inhibition of the psychiatric risk factor FKBP51 has anxiolytic properties. J Neurosci 2015;35:9007–16.
24. Holsboer F. The corticosteroid receptor hypothesis of depression. Neuropsychopharmacology 2000;23:477–501.
25. Howard DM, Adams MJ, Clarke TK, Hafferty JD, et al. Genome-wide meta-analysis of depression identifies 102 independent variants and highlights the importance of the prefrontal brain regions. Nat Neurosci 2019;22:343–52.
26. Israel-Elgali I, Hertzberg L, Shapira G, Segev A, et al. Blood transcriptional response to treatment-resistant depression during electroconvulsive therapy. J Psychiatr Res 2021;141:92–103.
27. Jagtap PKA, Asami S, Sippel C, Kaila VRI, et al. Selective inhibitors of FKBP51 employ conformational selection of dynamic invisible states. Angew Chem Int Ed Engl 2019;58:9429–33.
28. Kolos JM, Voll AM, Bauder M, Hausch F. FKBP ligands – where we are and where to go? Front Pharmacol 2018;9:1425.
29. Kumar R, Moche M, Winblad B, Pavlov PF. Combined x-ray crystallography and computational modeling approach to investigate the Hsp90 C-terminal peptide binding to FKBP51. Sci Rep 2017;7:14288.
30. LeMaster DM, Hernandez G. Conformational dynamics in FKBP domains: relevance to molecular signaling and drug design. Curr Mol Pharmacol 2015;9:5–26.
31. Li H, Su P, Lai TK, Jiang A, et al. The glucocorticoid receptor-FKBP51 complex contributes to fear conditioning and posttraumatic stress disorder. J Clin Invest 2020;130:877–89.
32. Modell S, Yassouridis A, Huber J, Holsboer F. Corticosteroid receptor function is decreased in depressed patients. Neuroendocrinology 1997;65:216–22.
33. Pei H, Li L, Fridley BL, Jenkins GD, et al. FKBP51 affects cancer cell response to chemotherapy by negatively regulating Akt. Cancer Cell 2009;16:259–66.
34. Pöhlmann ML, Häusl AS, Harbich D, Balsevich G, et al. Pharmacological modulation of the psychiatric risk factor FKBP51 alters efficiency of common antidepressant drugs. Front Behav Neurosci 2018;12:262.
35. Rein T. FK506 binding protein 51 integrates pathways of adaptation: FKBP51 shapes the reactivity to environmental change. Bioessays 2016;38894–902.
36. Sabbagh JJ, Cordova RA, Zheng D, Criado-Marrero M, et al. Targeting the FKBP51/GR/Hsp90 complex to identify functionally relevant treatments for depression and PTSD. ACS Chem Biol 2018;13:2288–99.
37. Scheufler C, Brinker A, Bourenkov G, Pegoraro S, et al. Structure of TPR domain-peptide complexes: critical elements in the assembly of the Hsp70-Hsp90 multichaperone machine. Cell 2000;101:199–210.
38. Schmidt MV, Paez-Pereda M, Holsboer F, Hausch F. The prospect of FKBP51 as a drug target. ChemMedChem 2012;7:1351–9.
39. Schülke JP, Wochnik GM, Lang-Rollin I, Gassen NC, et al. Differential impact of tetratricopeptide repeat proteins on the steroid hormone receptors. PLoS One 2010;5:e11717.
40. Sinars CR, Cheung-Flynn J, Rimerman RA, Scammell JG, et al. Structure of the large FK506-binding protein FKBP51, an Hsp90-binding protein and a component of steroid receptor complexes. Proc Natl Acad Sci U S A 2003;100:868–73.
41. Storer CL, Dickey CA, Galigniana MD, Rein T, et al. FKBP51 and FKBP52 in signaling and disease. Trends Endocrinol Metab 2011;22:481–90.
42. Thareja A, Hughes H, Alvarez-Lorenzo C, Hakkarainen JJ, et al. Penetration enhancers for topical drug delivery to the ocular posterior segment – a systematic review. Pharmaceutics 2021;13:276.
43. Voll AM, Meyners C, Taubert MC, Bajaj T, et al. Macrocyclic FKBP51 ligands define a transient binding mode with enhanced selectivity. Angew Chem Int Ed Engl 2021;60:13257–63.
44. Wochnik GM, Rüegg J, Abel GA, Schmidt U, et al. FK506-binding proteins 51 and 52 differentially regulate dynein interaction and nuclear translocation of the glucocorticoid receptor in mammalian cells. J Biol Chem 2005;280:4609–16.
45. Wray NR, Ripke S, Mattheisen M, Trzaskowski M, et al. Genome-wide association analyses identify 44 risk variants and refine the genetic architecture of major depression. Nat Genet 2018;50:668–81.
46. Wu B, Li P, Liu Y, Lou Z, et al. 3D structure of human FK506-binding protein 52: implications for the assembly of the glucocorticoid receptor/Hsp90/immunophilin heterocomplex. Proc Natl Acad Sci U S A 2004;101:8348–53.
47. Yeh WC, Li TK, Bierer BE, McKnight SL. Identification and characterization of an immunophilin expressed during the clonal expansion phase of adipocyte differentiation. Proc Natl Acad Sci U S A 1995;92:11081–5.
48. Zeytuni N, Zarivach R. Structural and functional discussion of the tetra-trico-peptide repeat, a protein interaction module. Structure 2012;20:397–405.
49. Zgajnar NR, De Leo SA, Lotufo CM, Erlejman AG, et al. Biological actions of the Hsp90-binding immunophilins FKBP51 and FKBP52. Biomolecules 2019;9:E52.
50. Zou L, Peng Q, Wang P, Zhou B. Progress in research and application of HIV-1 TAT-derived cell-penetrating peptide. J Membr Biol 2017;250:115–22.
Prof. Dr. Theo Rein, Max-Planck-Institut für Psychiatrie, Abteilung Translationale Forschung in der Psychiatrie, Kraepelinstraße 10, 80804 München, E-Mail: theorein@psych.mpg.de
FKBP5 ligands: potential antidepressants and more?
The obvious need for improved therapeutic option for depression contrasts the lack of new antidepressants. The potential of drug development on the basis of the monoamine deficiency hypothesis appears largely exhausted. The stress protein FKBP51 emerged in the recent years as promising candidate for the targeted development of new antidepressants. Challenges arise from the nature of the binding site on FKBP51 and the multiple functions of the protein. At the same time, the latter might be a chance for the development of ligands of versatile use.
Key words: depression, FKBP5, antidepressants, drug development, FKBP51, stress-related diseases
Psychopharmakotherapie 2022; 29(03):78-83