Walter E. Müller, Frankfurt/M., Anne Eckert, Basel, Gunter P. Eckert, Gießen, Heidrun Fink, Berlin, Kristina Friedland, Erlangen, Robert Hörr, Karlsruhe, Ralf Ihl, Krefeld, Siegfried Kasper, Wien, und Hans-Jürgen Möller, München
Von der traditionellen chinesischen Medizin zum evidenzbasierten Antidementivum
Die Blätter des Ginkgo-Baums beziehungsweise aus diesen Blättern hergestellte pharmazeutische Präparationen haben eine lange Geschichte in der traditionellen chinesischen Medizin, allerdings haben sie dort nie eine größere Bedeutung gewonnen, sodass paradoxerweise heute in China eher die modernen Extrakt-Präparate eingesetzt werden als traditionelle Zubereitungen [22, 23]. Die moderne Geschichte von Ginkgo als Arzneimittel begann 1965 mit der Einführung eines Ginkgo-Blätterextrakts durch Dr. Willmar Schwabe, einen Pharmazeuten und Mediziner, der das auf der Basis dieses Extrakts entwickelte Fertigpräparat (Tropfen und Tabletten) unter dem Namen Tebonin® mit seiner gleichnamigen Firma in den Handel brachte. Indikationen waren hier periphere, besonders auch zentrale Durchblutungsstörungen mit dem Schwerpunkt der sogenannten „Zerebralsklerose“ oder „Hirnverkalkung“, die zur damaligen Zeit eine wichtige ätiopathogenetische Erklärung für kognitive Defizite bei älteren Patienten war. Weiterführende Forschung und die Erkenntnisse, dass die meisten Inhaltsstoffe positive, einige wenige auch negative Effekte hatten, führte letztlich zur Einführung des Spezialextrakts EGb761® (Kasten 1) in den 80er-Jahren, in dem Substanzen wie die Flavonoide, die Ginkgolide und Bilobalid angereichert und Substanzen wie die Ginkgolsäuren aufgrund möglicher unerwünschter Wirkungen abgereichert waren. Dieser standardisierte Extrakt wurde zusammen mit der französischen Firma Ipsen entwickelt, die parallel zum deutschen Produkt Tebonin® den gleichen Extrakt unter dem Handelsnamen Tanakan® einführte. Beide Präparate stellen heute den größten Teil aller weltweit therapeutisch eingesetzten Ginkgoextrakt-Präparate dar [18].
Wie erwähnt, waren die ursprünglichen Indikationen für EGb761® periphere und zentrale Durchblutungsstörungen, wobei die Verwendung von EGb761® bei peripheren Störungen wie Raynaud-Syndrom, Claudicatio intermittens und peripherer arterieller Verschlusskrankheit heute weitgehend verlassen ist, obwohl durchaus positive Placebo-kontrollierte Studien nach den damaligen Standards vorlagen [23]. Im Hinblick auf den Einsatz bei zerebralen Durchblutungsstörungen, damals unter dem Konzept „zerebrale Insuffizienz“ subsumiert, hat man schon in den ersten Jahren der Verwendung positive Effekte, besonders auch auf kognitive Defizite gesehen, sodass der therapeutische Einsatz bei altersbezogenen Hirnleistungsstörungen über die nächsten Jahrzehnte weiterentwickelt wurde und den sich mit den Jahren ändernden unterschiedlichen Diagnosebegriffen angepasst wurde.
Kasten 1. Eigenschaften des Spezialextrakts EGb761®
Ausgangsprodukt sind getrocknete Blätter des Baumes Ginkgo biloba, die mit einem Aceton-Wasser-Gemisch (60%, m/m) extrahiert werden.
Der Extrakt hat dann ein Droge/Extrakt-Verhältnis von 35–67:1, das heißt, 1 g Extrakt geht auf 35 bis 67 g getrocknete Blätter zurück.
Die Zusammensetzung des fertigen Extrakts beträgt:
- 22–27% Ginkgoflavonoide
- 5–7% Terpenlactone
- 2,8–3,4% Ginkgolide A, B und C
- 2,6–3,2 Bilobalid
- <5 ppm Ginkgolsäure
Der fertige Extrakt besteht damit zu über 30% aus für die Wirksamkeit relevanten Inhaltsstoffen.
Durch das Verfahren wird die relative Zusammensetzung der Inhaltsstoffe im Vergleich zum Ausgangsmaterial angereichert (Flavonoide, Terpenlactone) bzw. abgereichert (potenziell toxische Ginkgolsäuren). Daher der Begriff Spezialextrakt, der in Deutschland unter dem Handelsnamen Tebonin® erhältlich ist.
Daten nach Ude et al. [119] und Biber [14]
Über viele Jahre hielt sich besonders auch in der deutschen Psychiatrie das Konzept des hirnorganischen Psychosyndroms (HOPS), oft synonym mit Hirnleistungsstörungen benutzt (Kasten 2), das syndromal dem heutigen Demenzbegriff sehr ähnlich war. Hier wurden zwar hauptsächlich ältere Patienten, Patienten mit Alzheimer- bzw. vaskulärer Demenz erfasst, allerdings auch andere Demenzen und beispielsweise auch delirante Patienten. Wegen dieser Unschärfe wurde der Begriff HOPS später durch die unterschiedlichen Beschreibungsformeln der Demenz und seit 1994 mit der Einführung des DSM-IV durch die Demenz vom Alzheimertyp bzw. die vaskuläre Demenz und Mischformen ersetzt (Tab. 1). Betrachtet man auf der anderen Seite die typischen Störungsbilder im Rahmen des hirnorganischen Psychosyndroms, wo Störungen der kognitiven Leistungsfähigkeit und Affektivität im Vordergrund stehen, ist es nicht weiter verwunderlich, dass ungeachtet der diagnostischen Zuordnung Patienten mit Hirnleistungsstörungen im Alter sehr ähnlich von EGb761® im Sinne einer Verbesserung der kognitiven Leistungsfähigkeit und des affektiven Status profitieren, wie wir es heute von Patienten mit einer vaskulären oder neurodegenerativen Demenz kennen (Tab. 1). Damit können auch die älteren Studien mit EGb761® bei HOPS in diese sich ändernden diagnostischen Konzepte eingepasst werden [39, 45, 78]. Dies gilt auch für die dem alten Begriff Hirnleistungsstörungen nicht ganz weit entfernte neue Einordnung dieser Krankheitsbilder unter dem Oberbegriff „neurokognitive Störungen“ in der DSM-5, sodass in einer aktuelle Sekundäranalyse der wichtigsten Therapiestudien mit EGb761® bei Demenz mit unterschiedlichen diagnostischen Konzepten (Tab. 1) die meisten Patienten auch den unterschiedlichen Klassen von neurokognitiven Störungen im Sinne der DSM-5 zugeordnet werden konnten [44].
Kasten 2. HOPS – Hirnorganisches Psychosyndrom (Organic brain syndrome)
- Definition: unscharf (oft synonym: Hirnleistungsstörung, Demenz)
- Beschreibung
- Intellektuelle Beeinträchtigung (Abstraktion, Bewertung)
- Gedächtnisstörung (Arbeitsgedächtnis, episodisches Gedächtnis)
- Störung von Urteilsvermögen, Handlungsplanung
- Störung von Orientierung, Aufmerksamkeit, Konzentration
- Störungen der Affektivität (Reizbarkeit, depressive Verstimmung, Angst)
Die Daten beziehen sich auf die Übersichten von Epstein und Simon [32], Berger [13] und Kanowski et al. [58].
Tab. 1. Demenz-Definitionen im Wandel der Zeit
|
1980 |
DSM-III |
Primär degenerative Demenz vom Alzheimer-Typ (senil/präsenil =Alzheimer-Krankheit), Multi-Infarkt-Demenz |
|
1986 |
NINCDS-ADRDA |
Probable/possible Alzheimer’s disease (AD) |
|
1986 |
DSM-III-R |
Primär degenerative Demenz vom Alzheimer-Typ (senil/präsenil), Multi-Infarkt-Demenz |
|
1993 |
NINDS-AIREN |
Probable/possible vascular dementia (für Mischform: possible AD with cerebrovascular disease) |
|
1994 |
DSM-IV |
Demenz vom Alzheimer-Typ, vaskuläre Demenz Heute modifiziert: Familiäre Alzheimer-Krankheit (fAD=early-onset AD, EOAD) Sporadische Alzheimer-Krankheit (sAD=late-onset AD, LOAD) |
|
2013 |
DSM-5 |
Neurokognitive Störung (neurocognitive disorder) mild oder stark ausgeprägt, jeweils vom Alzheimer- bzw. vaskulären Typ |
DSM: Diagnostic and statistical manual of mental disorders; NINCDS-ADRDA: National Institute of Neurological and Communicative Disorders and Stroke/Alzheimer’s Disease and Related Disorders Assciation; NINDS-AIREN: National Institute of Neurological Disorders and Stroke/Association Internationale pour la Récherche et l‘Enseignement en Neurosciences
Zur Zeit der EGb761®-Einführung dominierte die sogenannte „zerebrale Insuffizienz“ mit einer gefäßbedingten zerebralen Durchblutungsstörung als pathogenetisches Prinzip für Hirnleistungsstörungen im Alter. Allerdings gab es schon damals eher keine Hinweise auf einen primären vaskulären Angriffspunkt von EGb761® [23]. Mit der Erkenntnis der autonomen Regulation des zerebralen Blutkreislaufs und der dadurch bedingten physiologischen Anpassung der Durchblutung an den Bedarf hat das Konzept der primären Verbesserung der zerebralen Durchblutung an Bedeutung verloren. In den Vordergrund rückten Konzepte der neuronalen Dysfunktion mit gestörtem Energiestoffwechsel, Einschränkungen der zerebralen Neurotransmission, der Neuroplastizität und des neuronalen Überlebens. Interessanterweise konnte die fortgesetzte und intensivierte, auch präklinische Forschung zu EGb761® in den letzten Jahrzehnten zeigen, dass genau hier, also im Bereich der neuronalen Dysfunktion, ein wesentliches Wirkungsprinzip von EGb761 liegt, sodass die lange zurückliegenden Untersuchungen über eine Verbesserung der zerebralen Durchblutung durch EGb761® [41] eher als Ausdruck einer Verbesserung der neuronalen Funktion zu sehen sind, die ihren erhöhten Energiebedarf über eine Zunahme der Durchblutung bedient [79, 81]. Dies erklärt auch, warum praktisch alle Studien mit EGb761®, die getrennt die therapeutische Wirkung bei degenerativen (Alzheimer) Demenzen und vaskulären Demenzen untersucht haben, keine unterschiedliches Ansprechen gesehen haben (z.B. [58], [69] als ältere Arbeiten bzw. [85] als neuere Arbeit), da EGb761® in beiden Fällen eher über eine verbesserte neuronale Funktion wirkt (siehe Abschnitte „Kumulative neuronale Schädigung …“ und „Mitochondrien als Target“). Dies schließt nicht aus, dass direkte durchblutungsfördernde Effekte unter anderem über eine Verbesserung der rheologischen Eigenschaften des Bluts eine gewisse Rolle spielen [23, 26].
Da Neuronen einen großen Teil ihres Energiebedarfs für die intra-zerebrale Kommunikation von Neuronen, Projektionen und neuronalen Netzwerken benötigen, wirken sich Verbesserungen in diesem Bereich besonders auf kognitive Funktionen und die Regulation der Gleichgewichtskontrolle aus, weil gerade diese zerebralen Funktionen auf über viele Hirnareale gehende Netzwerkstrukturen angewiesen sind und daher besonders empfindlich auf eine Einschränkung der Energiebereitstellung reagieren. Die klinische Wirksamkeit von EGb761® bei Hirnleistungsstörungen im Alter über deren gesamte Breite (s.u.) und bei Störungen der Gleichgewichtskontrolle (Schwindel) [40, 108], besonders bei älteren Patienten, lässt sich daraus erklären. EGb761® ist damit kein Neuroenhancer, denn junge Gesunde profitieren wenig, wenn überhaupt von Ginkgo, sondern es ist eine Substanz, die neuronale Funktion gezielt im Falle einer funktionellen Beeinträchtigung verbessert und damit auch die daraus resultierenden kognitiven und emotionale Einschränkungen.
Kognitionsverbesserung im Tierversuch: Verbesserung vorbestehender Defizite steht im Vordergrund
Wie ausgeführt, hat sich über die letzten 20 bis 30 Jahre das Konzept herauskristallisiert, dass EGb761® kognitive Verbesserungen eher nur dann zeigt, wenn vorbestehende, durch unterschiedliche Noxen ausgelöste Defizite existieren, zum Beispiel unterschiedliche Formen von alterseingeschränkter kognitiver Leistungsfähigkeit. Es besteht damit kein unspezifisches Neuroenhancement, sondern eine Gegensteuerung von kognitiven Einschränkungen, die mit neuronaler Dysfunktion auf unterschiedlichen Ebenen einhergehen.
Dieses Konzept gilt nicht nur für die klinische Literatur zur EGb761®, sondern zeichnet sich auch in den tierexperimentellen Untersuchungen mit EGb761® im Hinblick auf kognitive Verbesserungen ab. Wichtige, meist neuere, ab etwa 1990 publizierte tierexperimentelle Arbeiten sind in Tabelle 2 zusammengefasst. Eine ältere Zusammenfassung ist die Übersicht von Müller und Chatterjee [80].
Tab. 2. Effekte von EGb761® auf die Kognition im Tierversuch
|
EGb761®-Dosierung |
Tiermodell |
Verhaltenstest |
Wirkung |
Literatur |
|
p.o. 5 Monate |
TgCRND8 APP-transgene Maus |
Barnes maze |
+ |
Liu et al. [74] |
|
p.o. 21 d |
Restraint Stress/Corticosteron Ratte |
Morris water maze Radial arm maze |
+ + |
Walesiuk & Braszko [125] |
|
p.o. 3 Monate |
Alte Ratte |
Morris water maze |
+ |
Blecharz-Klin et al. [15] |
|
i.p. 10 d/14 d/20 d |
Ratte |
Double Y maze/-pretraining Double Y maze/-postraining |
+ Ø |
Satvat & Mallet [98] |
|
p.o. 1 d p.o. 7 d |
Ratte |
Fear conditioning |
(+) |
Oliveira et al. [87] |
|
p.o. 87 d |
Ovarektomie + Restraint Stress Ratte |
Novel object recognition |
+ |
Takuma et al. [115] |
|
p.o. 21 d |
Restraint Stress/Corticosteron Ratte |
Passive avoidance Conditioned avoidance |
+ Ø |
Walesiuk et al. [124] |
|
p.o. 30 d |
Alte Ratte |
Morris water maze Platform switched Morris water maze |
+ + |
Wang et al. [127] |
|
p.o. 4 d |
Transiente, globale zerebrale Ischämie Ratte |
Non-food motivated, aversive radial arm maze |
+ |
Paganelli et al. [88] |
|
p.o. 21 d |
Restraint Stress/Corticosteron Ratte |
Morris water maze Novel object recognition |
+ + |
Walesiuk et al. [123] |
|
p.o. 7 Monate |
Chronischer, ultramilder Stress Alte Maus |
Decision making (T maze) |
+ |
Pardon et al. [89] |
|
p.o. 80 d |
Chronische, bilaterale A.-carotis-Ligatur Ratte |
Radial arm maze |
+ |
Lin et al. [73] |
|
i.p. 30 d/1 d |
Adulte Ratte/Alte Ratte |
Olfactory recognition |
+ |
Wirth et al. [135] |
|
p.o. 70 d |
i. c. v. Streptozotocin Ratte |
Holeboard Passive avoidance |
+ + |
Hoyer et al. [47] |
|
p.o. >24 Monate |
Alte Ratte |
Radial arm maze |
+ |
Winter JC [134] |
|
i.p. 7 d |
Bilateral frontale Kortex-Kontusion Ratte |
Morris water maze |
+ |
Hoffman & Stein [46] |
|
21 d |
Junge, adulte & alte Maus |
Passive avoidance |
+ |
Stoll et al. [113] |
|
p.o. 3,5 Monate p.o. 4,5 Monate |
Maus |
Operant conditioning |
+ |
Winter E [133] |
|
p.o. 1–5 d |
Ratte |
Learned helplessness Passive avoidance |
+ Ø |
Porsolt et al. [93] |
|
Maus |
Four plates |
Ø |
+ Signifikante Verbesserung im Vergleich zur Kontrolle; Ø kein Unterschied zur Kontrolle
Wesentliche gemeinsame Aspekte aller dieser Arbeiten sind eine Behandlung mit EGb761® über mindestens ein bis zwei Wochen und der Befund, dass positive Effekte auf kognitive Parameter besonders dann beobachtet wurden, wenn vorbestehende Einschränkungen existierten, wobei kognitive Einschränkungen entweder über den Alterungsprozess, über experimentelle Infarktmodelle, chronischen Stress oder/und hormonelle Eingriffe ausgelöst wurden (Tab. 2). In einem transgenen Alzheimer-Mausmodell konnten ganz aktuell ähnliche Effekte gezeigt werden, nämlich eine Verbesserung des räumlichen Lernens bei den transgenen „Alzheimer“-Tieren [74]. Eine weitere aktuelle Arbeit konnte im Morris-Water-Maze noch einmal bestätigen, dass alte, aber nicht junge Tiere (Ratten) nach 30-tägiger Behandlung mit EGb761® eine kognitive Verbesserung im Morris-Water-Maze parallel zu einer Reduktion von oxidativem Stress zeigten [12].
Man kann daher heute davon ausgehen, dass sich die kognitionsverbessernden Effekte von EGb761® im Tiermodell verifizieren lassen. Sie sind besonders ausgeprägt nach unterschiedlichen funktionellen Störungen des zentralen Nervensystems und beinhalten kognitive Domänen wie Arbeitsgedächtnis, räumliches Gedächtnis und Lernvermögen (Tab. 2).
Kumulative neuronale Schädigung als Ursache von Hirnleistungsstörungen im Alter und als therapeutisches Target von EGb761®
Pathophysiologische Grundlagen
Die früheren Konzepte einer klaren Trennung der biologischen Ursachen für leichtere kognitive Störungen im Rahmen des unspezifischen Hirnalterungsprozesses auf der einen Seite und den ausgeprägten Störungen der eigentlichen Demenzen, besonders der Demenz vom Alzheimer-Typ, auf der anderen Seite ist nach heutigen Vorstellungen so nicht mehr haltbar. Man geht vielmehr von einem Kontinuum neuronaler Schädigungen mit ähnlichen Ursachen im Frühstadium und ihrer Akzentuierung im Endstadium aus, allerdings mit deutlichen Unterschieden besonders im Hinblick auf das Ausmaß des Substanzverlusts (Verlust von Synapsen, Neuriten und letztlich von Nervenzellen durch Apoptose) [95].
Das ursprüngliche Konzept, die sogenannte Amyloid-Kaskaden-Hypothese, hatte relativ einseitig postuliert, dass die vermehrte Bildung des kleinen Beta-Amyloid-Peptids (Aβ) und seine Ablagerung in Form der extrazellulären Plaques den kritischen Prozess ausmacht, der die gesamte Alzheimer-Erkrankung letztlich initiiert und aufrecht erhält. Dies ist heute so nicht mehr haltbar. Wir wissen durch moderne Bildgebung, dass im Fall der späten, sporadischen Alzheimer-Erkrankung (LOAD, Tab. 1) Amyloid-haltige Plaques Jahrzehnte vor den ersten kognitiven Symptomen auftreten, sehr lange in ihrer Konzentration stabil sind und dann erstaunlicherweise sogar parallel zum Auftreten erster kognitiver Beeinträchtigungen eher ab- als weiter zunehmen [110, 114]. Auch bei den beiden genetischen Formen der Erkrankung, wo die Beobachtung, dass Mutationen der Risikogene (APP und PS1 bzw. PS2) zu massiver Zunahme der Aβ-Plaques führen, ein wichtiges Argument für die Amyloid-Kaskaden-Hypothese darstellte, gibt es Befunde, die dies infrage stellen, beispielsweise Daten, dass PS-1-Mutationen direkt, also Aβ-unabhängig Neurodegeneration auslösen können [8, 10, 17, 104]. Daher ist es nicht verwunderlich, dass sämtliche Therapieansätze, die Alzheimer-Erkrankung über eine Reduktion der Amyloid-haltigen Plaques zu therapieren, gänzlich fehlgeschlagen sind [52, 59, 114], obwohl in den meisten Fällen eine Reduktion der Amyloid-haltigen Plaques erreicht wurde.
Als Modell zur Erklärung der sporadischen Alzheimer-Erkrankung tritt daher die mitochondriale Kaskadenhypothese immer mehr in den Vordergrund (Abb. 1). Sie geht davon aus, dass bei Risikopersonen eine individuell hohe Belastung mit oxidativem Stress im Wesentlichen durch mitochondriale Dysfunktion über Jahre kumuliert, um dann einen kritischen Wert zu erreichen, wo sich diese Schädigungen auch über vermehrte Bildung von löslichen Beta-Amyloid-Oligomeren intensivieren und dann zu den typischen histologischen Schäden führen (Synapsenverlust und weitere Aspekte einer Neurodegeneration), die in leichter Ausprägung sehr viel früher auftreten als die typischen Alzheimer-Veränderungen (Plaques und Tangles). Oxidativer Stress entsteht immer dann, wenn das Gleichgewicht zwischen der Bildung freier Radikale, besonders von reaktiven Sauerstoffspezies (ROS), und ihrer Entgiftung durch bestimmte Enzyme wie SOD gestört ist (Abb. 2). Wesentliche Ursache ist in den meisten Fällen eine vermehrte Bildung von ROS, wofür mit zunehmendem Alter gestörte mitochondriale Funktion eine dominierende Rolle spielt. Diese kann initial durch eine kompensatorische Erhöhung der entgiftenden Enzyme (Abb. 2) kompensiert werden; ab einem gewissen Punkt reicht dies allerdings nicht mehr aus, sodass es zur vermehrten Schädigung von Gewebestrukturen durch die dann freien Radikale (ROS) kommt (Abb. 2). Mitochondriale Störungen spielen hier eine dominierende Rolle, da durch die Bildung von ROS in den Mitochondrien die Mitochondrien auch direkt geschädigt werden. Der gesamte Prozess kann sich erheblich selbst verstärken, Folgen sind eine Störung bzw. Schädigung der neuronalen Funktion durch eine reduzierte Verfügbarkeit von ATP als zellulärem Energieträger, besonders ausgeprägt an der Synapse als wesentlichem Energieverbraucher, und in der finalen Konsequenz ein Absterben der Nervenzelle durch Apoptose (Abb. 2).
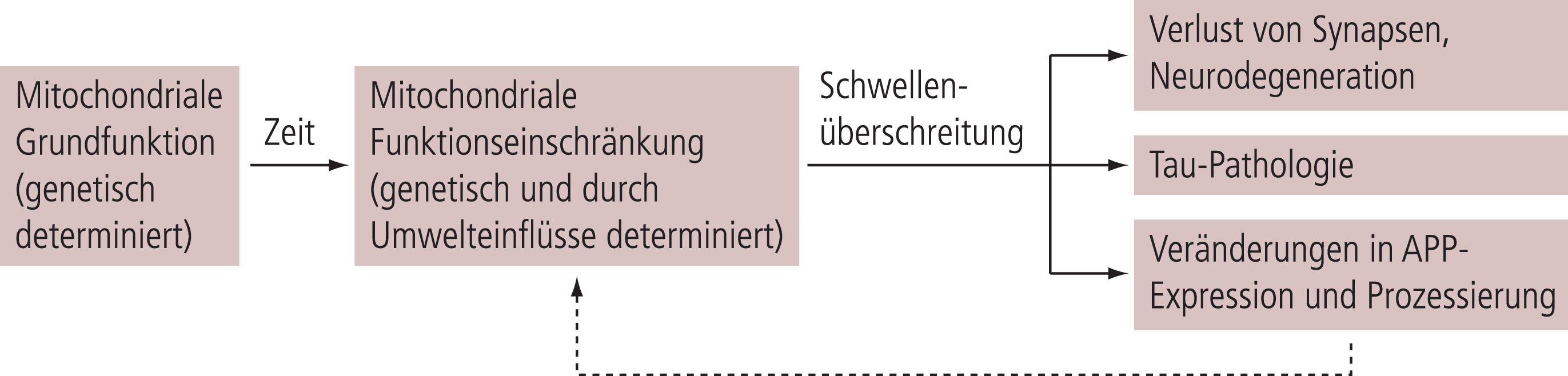
Abb. 1. Mitochondriale Kaskaden-Hypothese. Die aktuelle mitochondriale Kaskaden-Hypothese postuliert, dass eine individuelle Beeinträchtigung der Mitochondrienfunktion (z.B. genetisch determiniert) ein wesentlicher Risikofaktor für eine spätere Alzheimer-Erkrankung ist. Weiterhin durch individuelle genetische und Umweltfaktoren determiniert, nimmt die Dysfunktion über Jahrzehnte zu, um dann einen Schwellenwert zu erreichen, wo vermehrt noch lösliche neurotoxische Amyloid-beta(Aβ)-Oligomere gebildet werden, Tau-Pathologie und synaptische Dysfunktion starten gefolgt von kognitiven Beeinträchtigungen. Aβ ist hier nur einer der involvierten Faktoren, der allerdings erst später ins Spiel kommt (nach Swertlow et al. [114]). Wichtige für die Hypothese relevante genetische Risikofaktoren sind APOE4 und PS1, die auch Aβ-unabhängig oxidativen Stress verstärken können [19, 48, 104]. APP: Amyloid-Precursor-Protein; APOE: Apolipoprotein E; PS: Präsenilin
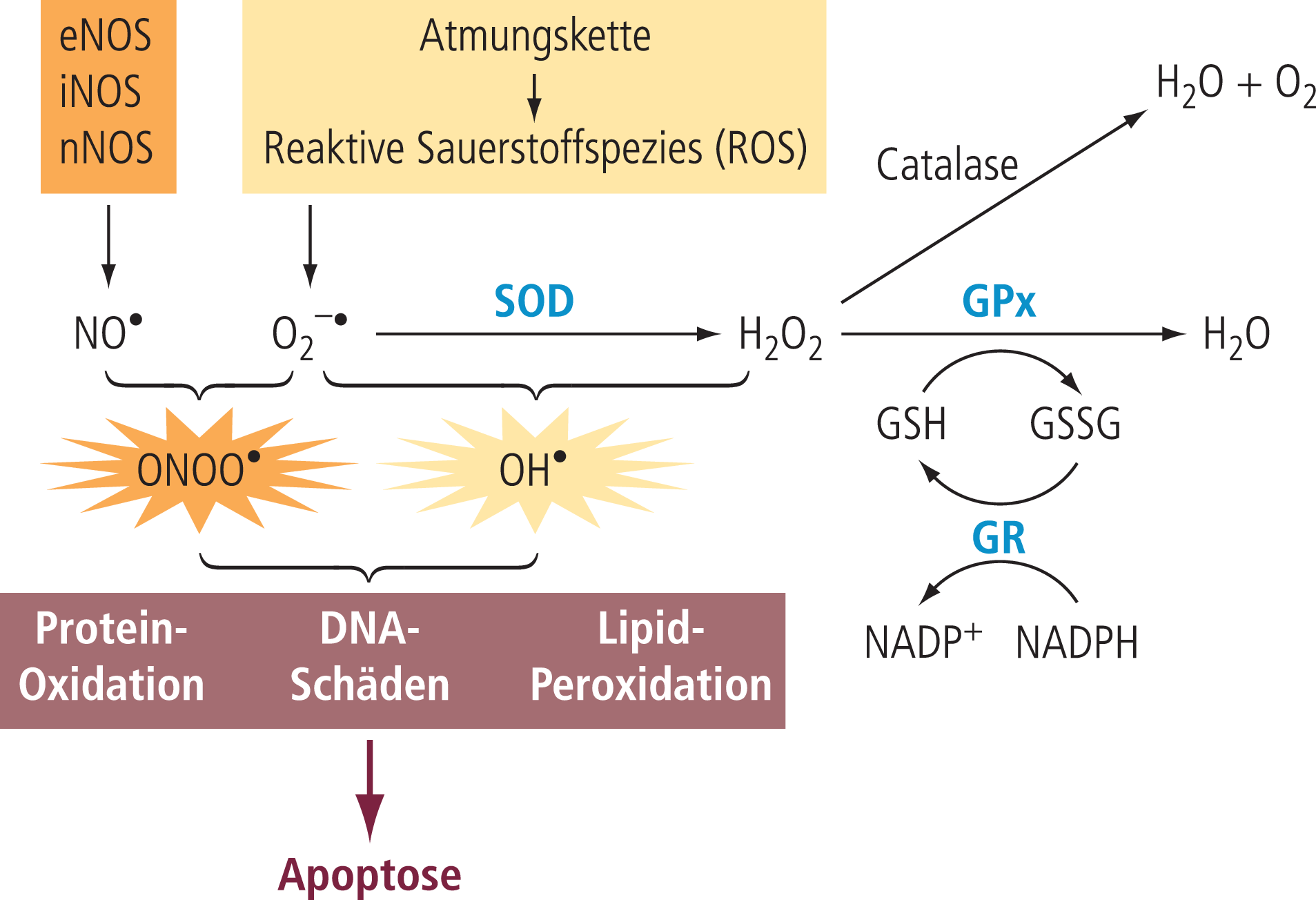
Abb. 2. Oxidativer Stress, ein wichtiger Grund für Hirnalterungsprozesse: Oxidativer Stress, eine Zunahme von gewebeschädigenden freien Radikalen (primär Sauerstoff), entsteht hauptsächlich durch die Atmungskette, wenn zu viel ROS gebildet wird und/oder die entgiftenden Enzyme die Radikale nicht schnell genug abbauen können. Folge sind Gewebeschäden, besonders auch am Bildungsort der Mitochondrien. G: Glutathion; GPx: Glutathionperoxidase; GR: Glutathionreductase; SOD: Superoxiddismutase
Wenn mit zunehmendem Alter (Abb. 1) eine vermehrte ROS-Konzentration über eine direkte Stimulation der Gamma-Secretase [71, 72] die Bildung von Beta-Amyloid massiv erhöht (Abb. 3), entsteht darüber hinaus ein Szenarium, in dem sich Alterseffekt und Beta-Amyloid-Toxizität (noch lösliche Oligomere) gegenseitig hochspielen [112].
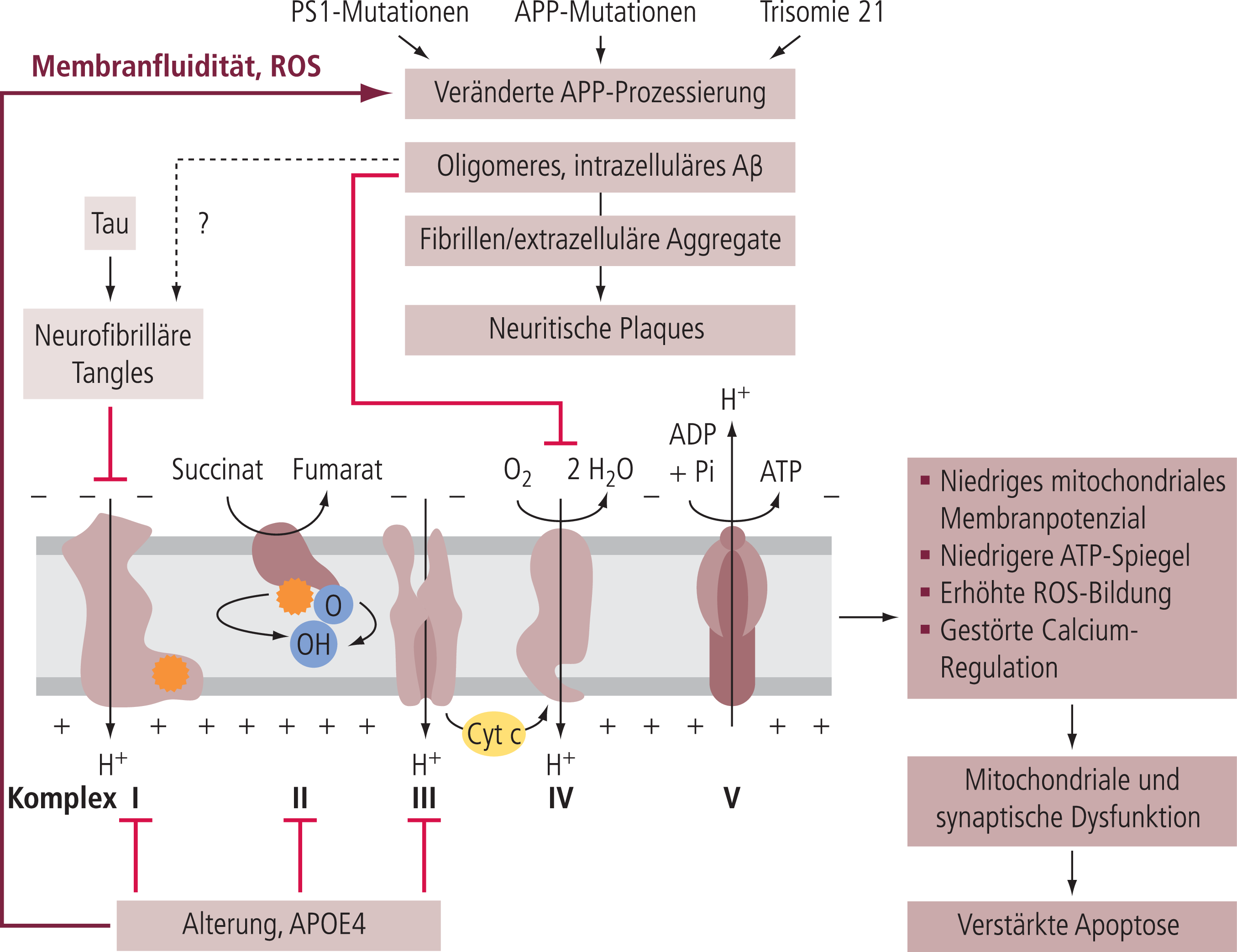
Abb. 3. Mitochondriale Dysfunktion als gemeinsame Endstrecke spezifischer und unspezifischer Risikofaktoren der Alzheimer-Demenz. Altern, Tau-Tangles und Beta-Amyloid-Plaques führen zu einer verringerten Aktivität von unterschiedlichen Atmungsketten-Komplexen und somit zu einem verringerten mitochondrialen Membranpotenzial, zu reduziertem ATP-Spiegeln und zu einer erhöhten Produktion an reaktiven Sauerstoffspezies (ROS). Dies mündet in einer mitochondrialen und synaptischen Dysfunktion, welche wiederum zum Zelltod führt. Das durch genetische Risikofaktoren bzw. den Alterungsprozess vermehrt gebildete Aβ kann dann wiederum vermehrt zu ROS-Bildung führen, sodass die Prozesse sich im Sinne eines Teufelskreises gegenseitig verstärken können [70, 71, 72, 83].
Kumulative neuronale Schädigung als spezifisches Target von EGb761®
Im Rahmen des beschriebenen Konzepts zur Pathophysiologie altersbedingter Hirnleistungsstörungen müssen heute die pharmakologischen Eigenschaften von EGb761® eingeordnet werden, die auf den ersten Blick relativ unzusammenhängend erscheinen, sich letztlich aber auf wenige primäre Mechanismen reduzieren lassen. Die wesentlichen Eigenschaften können wie folgt zusammengefasst werden [77, 79, 106]:
- Direkte antioxidative radikalfangende Eigenschaften
- Mitochondriale Protektion bzw. Verbesserung der mitochondrialen Funktion
- Neurotrophe Eigenschaften und Verbesserung der synaptischen Plastizität
- Verbesserung der Neurotransmission
- Hemmung der Thrombozytenaggregation und Verbesserung der Erythrozytenverformbarkeit
(1) Die radikalfangenden bzw. antioxidativen Eigenschaften gehen besonders auf die Flavonoidfraktion zurück. Flavonoide bzw. Polyphenole sind nicht spezifisch für Ginkgo-Extrakte wie EGb761®, sondern kommen in sehr vielen Pflanzen vor, besonders auch in Gemüse und Obst. Damit können antioxidative Eigenschaften nicht allein das breite klinische Wirksamkeitsspektrum von EGb761® erklären, aber man kann mit Sicherheit annehmen, dass sie zu mindestens auch an der Langzeitwirkung bei Hirnleistungsstörungen beteiligt sind. Dafür spricht unter anderem eine Reihe epidemiologischer Daten, die darauf hinweisen, dass hoher Konsum von Flavonoid-haltigen Lebensmitteln mit einer Senkung des Demenzrisikos verbunden zu sein scheint [21, 25, 31].
(2) Mitochondriale Protektion bzw. Verbesserung der mitochondrialen Funktion scheint ein zentraler Wirkungsmechanismus aller EGb761®-Bestandteile zu sein, wobei allerdings die Flavonoide weniger beteiligt zu sein scheinen als die Isoterpenlactone wie Ginkgolide und Bilobalid, die hier einen direkten, spezifischen Effekt zeigen. Aufgrund der großen Bedeutung dieses Mechanismus für die Wirksamkeit von EGb761® wird dieser Aspekt in einem gesonderten Absatz („Mitochondrien als Target“) ausführlich besprochen. Wichtig ist nur an dieser Stelle zu erwähnen, dass durch eine verbesserte mitochondriale Funktion der neuronalen Zelle mehr Energie in Form von ATP bereitgestellt wird, was wiederum zu einer Zunahme der neuronalen Funktion führt; das Neuron benötigt dann auch mehr Sauerstoff, der letztlich über die Autoregulation der Hirndurchblutung über eine Zunahme der Durchblutung herangeführt wird. Eine verbesserte Energiesituation der Zelle, besonders im Bereich der Synapsen, wo sehr viel Energie verbraucht wird, kann eine verbesserte präsynaptische und postsynaptische Funktion [33, 65, 139] bewirken.
(3) Eine Zunahme der neuronalen bzw. synaptischen Plastizität unter EGb761 ist sicher nicht alleine Ausdruck des verbesserten Energiestoffwechsels über eine verbesserte mitochondriale Funktion, sondern alle Bestandteile von Ginkgo, hier auch wieder die Flavonoide, können offensichtlich in die Signalkaskaden eingreifen, die die Steuerung der komplexen Mechanismen zur Optimierung der synaptischen Plastizität auf allen Ebenen bedingen. Dieser wichtige Punkt wird im Kapitel „Effekt auf die Neuroplastizität“ noch einmal ausführlich dargestellt.
(4) Eine Verbesserung der chemischen Neurotransmission ist in verschiedenen Untersuchungen für verschiedene Neurotransmitter, besonders aber für die biogenen Amine Noradrenalin, Serotonin und Dopamin gezeigt worden. Weil einige dieser Effekte möglicherweise auch über den verbesserten Energiestoffwechsel erklärt werden können, scheinen aber auch für einzelne EGb761®-Bestandteile direkte Effekte auf Mechanismen der Neurotransmission, beispielsweise die neuronale Rückaufnahme bzw. auch die neuronale Freisetzung der Aminneurotransmitter, eine Rolle zu spielen [3, 65, 139]. Der Mechanismus dieser Effekte ist nicht sicher bekannt, allerdings konnte gezeigt werden, dass verschiedene Bestandteile von EGb761 in unterschiedlichem Maß direkt die Aufnahmeprozesse von Noradrenalin, Serotonin und Dopamin in isolierten Nervenendigungen hemmen können [33].
(5) Einige Ginkgolide sind Antagonisten des Plättchenaktivierungsfaktors (PAF). Über PAF-Antagonismus können diese Bestandteile von EGb761® die Thrombozytenaggregation etwas reduzieren und damit zusammen mit einem verbessernden Effekt auf die Erythrozytenverformbarkeit über eine verbesserte rheologische Situation des Bluts zu einer geringfügig verbesserten Durchblutung beitragen [26]. Rein theoretisch könnte dieser Mechanismus auch bei therapeutischer Anwendung zu einer Senkung der Gerinnungsfähigkeit des Bluts führen. Dies wird zwar in den Fachinformationen zu EGb761® aufgeführt, ist wissenschaftlich aber nicht belegt, da alle zur Verfügung stehenden klinischen Daten keine Hinweise ergeben haben, dass die Einnahme von EGb761® mit einem erhöhten Blutungsrisiko verbunden ist [64, 66]. Die Erklärung liegt wahrscheinlich darin, dass unter therapeutischen Bedingungen Plasmaspiegel der Ginkgolide erreicht werden, die weit unter den Konzentrationen liegen, die für einen PAF-Antagonismus benötigt werden [67].
Mitochondrien als Target
Die Akzeptanz von EGb761® als eine der Therapieoptionen für die Alzheimer-Erkrankung hat in den zurückliegenden Jahren darunter gelitten, dass die lange dominierende Beta-Amyloid-Kaskaden-Hypothese nicht primär von EGb761 bedient wird, da zwar im Einzelfall auch Reduktionen der Aβ-Produktion im zellulären Modell oder im Tiermodell gezeigt werden konnten [96], diese Effekte aber deutlich geringer waren als Reduktionen der Aβ-Belastung durch spezifische Aβ-gerichtete Substanzen wie Gamma-Secretasehemmer [20]. Die sehr deutlichen, lange bekannten Effekte von EGb761® auf mitochondriale Dysfunktion [3, 29, 30, 47, 53, 56, 96, 97] wurden als eher unspezifisch gesehen und konnten zunächst keine Akzeptanz als Erklärung für die klinische Wirksamkeit finden. Erst die Erkenntnis, dass eine mitochondriale Dysfunktion im Zentrum der sich über Jahrzehnte entwickelnden Pathologie der Alzheimer-Erkrankung steht, hat dazu geführt, dass die mitochondriale Wirkung von EGb761® die Substanz in das Zentrum unserer Konzepte zum pathophysiologischen Geschehens der Erkrankung katapultierte.
Warum sind Mitochondrien für eine neurodegenerative Erkrankung so wichtig? Die Antwort ergibt sich sehr einfach aus der Funktion der Mitochondrien, die zum einen durch Bereitstellung von ATP die Zellfunktion aufrecht halten und im Falle einer extremen Dysfunktion den Suizid (Apoptose) einleiten, um die anderen Zellen zu schützen (Abb. 2 und 4).
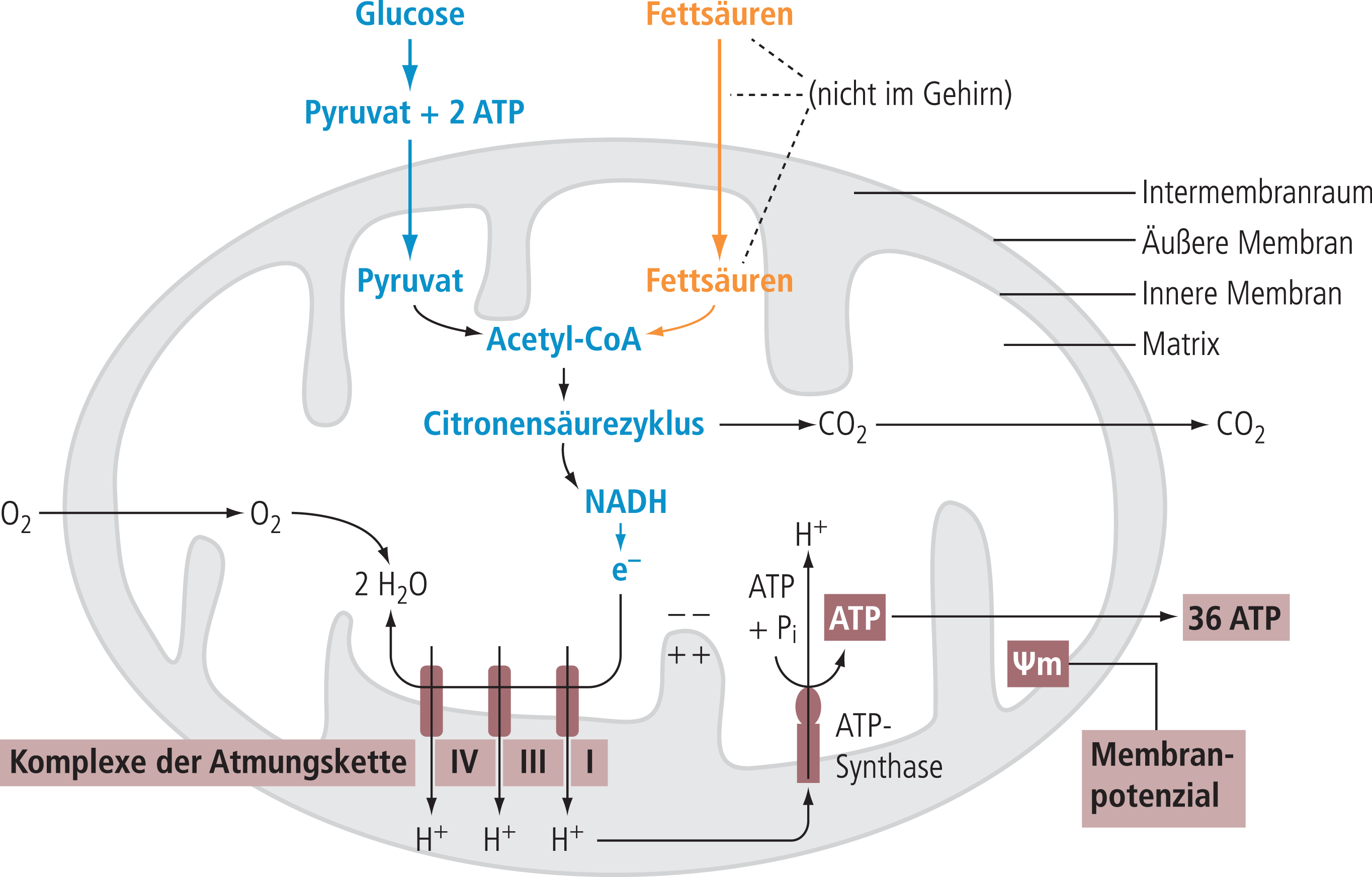
Abb. 4. Organisation und Funktion der Mitochondrien als Target für EGb761®. Mitochondrien produzieren über die Atmungskette Protonen (H+), die einen elektrischen Gradienten zwischen der inneren und äußeren Membran aufbauen, der letztlich die ATP-Produktion treibt. EGb761® verbessert bei gestörter mitochondrialer Funktion verschiedene Mechanismen der mitochondrialen Funktion (unterlegt) wie das Membranpotenzial, die Funktion der einzelnen Komplexe der Atmungskette und die ATP-Produktion und reduziert das Abgleiten in die Apoptose, als letzten Ausweg der Zelle bei irreparabler Störung der mitochondrialen Funktion.
Sehr früh hat man schon zeigen können, dass EGb761®, besonders aber auch die Bestandteile Bilobalid und die Ginkgolide, durch oxidativen Stress induzierten, aber auch durch andere Trigger wie Beta-Amyloid ausgelösten apoptotischen Zelltod in einer Reihe von Zellmodellen reduzieren konnten, zum Teil schon bei sehr niedrigen Konzentrationen der einzelnen Bestandteile [4, 11]. Auch Ex-vivo-Untersuchungen an Ratten (Behandlung der Tiere mit folgender Isolierung der Zellen nach der Tötung) konnten hier eine deutliche anti-apoptotische Wirkung zeigen [101].
Nachfolgende Untersuchungen haben dann gezeigt, dass EGb761® über weite Bereiche der mitochondrialen Funktion positive Effekte zeigt [70, 82]. EGb761® scheint eine protektive bzw. verbessernde Wirkung nicht nur durch die radikalfangenden Eigenschaften, sondern auch durch protektive Effekte an den einzelnen Komplexen der mitochondrialen Atmungskette zu besitzen (Abb. 4) [3, 9]. Andere Befunde zeigen in vitro, aber auch in Ex-vivo-Untersuchungen eine Verbesserung des gestörten mitochondrialen Membranpotenzials durch EGb761® bzw. eine Zunahme der ATP-Produktion unter ähnlichen Bedingungen (Abb. 4). Auch Messungen der oxidativen Phosphorylierung (Oxphos-Aktivität) an isolierten Zellen bzw. an isolierten Mitochondrien konnten eine Zunahme der Atmungskettenaktivität mit verbesserter ATP-Produktion unter der Einwirkung von Ginkgo-Extrakt zeigen, wobei letztere Ex-vivo-Versuche auch belegen, dass die Mitochondrien direkt von Ginkgo-Extrakt moduliert werden. Ursprüngliche Befunde, dass unter den einzelnen Ginkgo-Komponenten besonders Komplex I, Komplex III und Komplex IV Funktionsverbesserungen zeigen [70], gewonnen an isolierten Zellen der Maus, konnten in jüngster Zeit an SY5Y-Zellen in vitro bestätigt werden, wo darüber hinaus gezeigt werden konnte, dass es zu einer Zunahme der Mitochondrien-Biosynthese unter der Einwirkung von Ginkgo kommt [96]. Die Verbesserung der mitochondrialen Funktion und die damit verbundene Produktion von ROS können auch die bereits erwähnten Effekte von Ginkgo und die Aβ-Produktion erklären (Abb. 3).
Zusammenfassend ist die Datenlage, die hier nur kursorisch wiedergegeben werden konnte dahingehend sehr überzeugend, dass EGb761® bzw. alle seine drei wirksamen Komponenten die Funktion von Mitochondrien verbessern, besonders dann, wenn durch verschiedene Noxen, besonders aber durch oxidativen Stress im weitesten Sinne die mitochondriale Funktion beeinträchtigt ist. Dies deckt sich sehr gut mit den aktuellen Vorstellungen über eine breite Bedeutung mitochondrialer Störungen für das gesamte Kontinuum von Hirnleistungsstörungen im Alter, von leichteren, eher altersbedingten Veränderungen hin bis zu Vorstufen der Alzheimer-Demenz wie MCI (Minimal cognitive impairment) bis hin zum Vollbild einer demenziellen Störung im Rahmen der Alzheimer-Erkrankung oder auch vaskulärer Störungsbilder.
Effekte auf die Neuroplastizität
Als neurodegenerative Demenz ist die Alzheimer-Erkrankung durch massive neuronale Schädigungen gekennzeichnet, wobei das Absterben von Neuronen eher im späteren Stadium auftritt, während Defekte von Synapsen und Neuriten schon eher in frühen Stadien auftreten und zudem im Rahmen des gesamten Kontinuums von Hirnleistungsstörungen, also auch schon im Alterungsprozess, nachweisbar sind [99, 100]. Im Gegensatz zu den Nervenzellen, die mit Ausnahme zweier kleinerer hippocampaler Bereiche, die Neurogenese zeigen, statisch sind, das heißt, nach Reifung des Gehirns bis zum Absterben ihre Zahl nicht verändern, unterliegen die synaptischen Strukturen einer ausgesprochenen Plastizität, das heißt, Neuriten und postsynaptischen Spines unterliegen einem konstanten Auf- und Abbau, um sich dem Bedarf anzupassen. Dass dieser Prozess viel Energie benötigt und daher bei mitochondrialer Dysfunktion, wie für das Kontinuum der Hirnleistungsstörungen typisch, gestört ist, liegt auf der Hand.
Synaptische Plastizität umfasst alle Bereiche der synaptischen Funktion, von der Ebene der eigentlichen Signaltransduktion (Rezeptoren, Transporter, Enzyme), der Ebene der Signalimprägnierung (hier besonders die Langzeit-Potenzierung) bis zu der für die Hirnalterung besonders wichtigen strukturellen (histologischen) Ebene mit Spine-Struktur und Spine-Dichte, Neuritogenese und Neurogenese (Abb. 5). Über die ganze Bandbreite der Mechanismen der synaptischen Plastizität konnten verbessernde Effekte von EGb761® gesehen werden (Tab. 3; [81]). Da in vielen Modellen die Effekte besonders ausgeprägt waren, wenn die Neuroplastizität durch Alter oder AD-analoge Veränderungen reduziert war, kann man davon ausgehen, dass eine verbesserte mitochondriale Funktion durch Ginkgo gefolgt von einer verbesserten ATP-Bereitstellung eine wichtige Rolle spielt. Auf der anderen Seite können verschiedene Ginkgo-Inhaltsstoffe auch direkt die Neuroplastizität verbessern, wahrscheinlich über einen Eingriff in die intrazellulären Signalkaskaden, die pCREB (Cyclic-AMP response element binding protein) aktivieren, einen Transkriptionsfaktor für viele neurotrophe Signalmoleküle [117, 118]. Auch unsere Daten über eine direkte Stimulation der Neuritogenese durch EGb761® bzw. einige wichtige Inhaltsstoffe sprechen für einen direkten Effekt auf die Signalmechanismen [81].
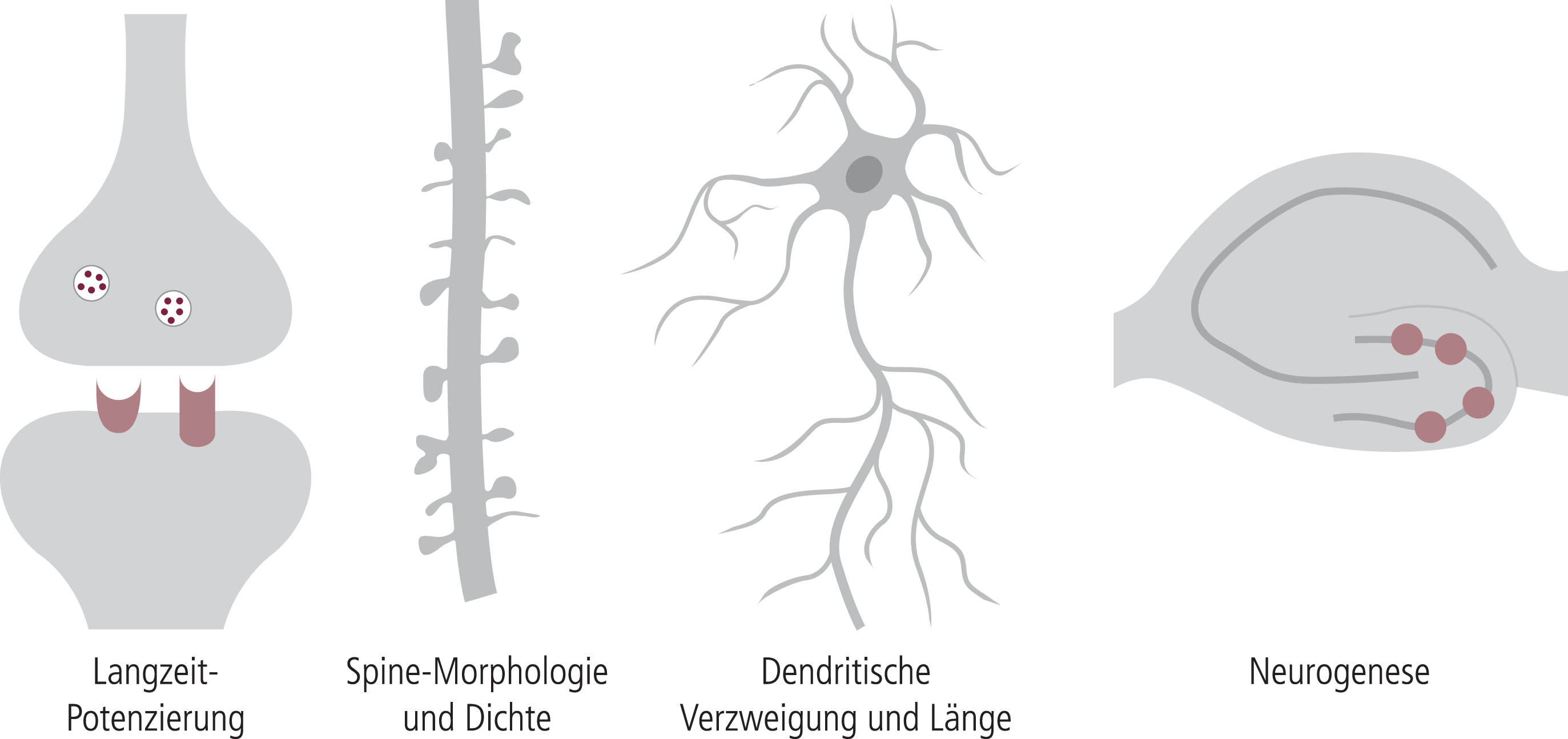
Abb. 5. Verschiedene adaptive Prozesse werden unter dem Begriff „neuronale (synaptische) Plastizität“ zusammengefasst
Tab. 3. Effekte von Ginkgo-Extrakt auf die verschiedenen Aspekte der Neuroplastizität an isolierten Zellen oder im Tierversuch
|
Langzeit-Potenzierung (LTP) |
|
|
Williams et al. [131] |
Langzeit-Potenzierung ist reduziert im Hippocampus alter Mäuse. EGb761®-Behandlung (300 mg/kg) verbesserte LTP im Hippocampus der alten Tiere, hatte keinen Effekt in den jungen Tieren. |
|
Vitolo et al. [121] |
Aβ1–42 hemmt LTP in Hippocampus-Schnitten von Mäusen. Ginkgo-Extrakt antagonisiert diesen Effekt. |
|
Wang et al. [127] |
LTP im Hippocampus und räumliches Gedächtnis sind in alten Ratten reduziert. Beide Parameter profitieren von EGb761®-Behandlung nur bei den alten Tieren. |
|
Spine-Morphologie |
|
|
Tchantchou et al. [118] |
Erhöhte Spine-Dichte an mit Aβ behandelten hippocampalen Neuronen durch Bilobalid und Quercetin |
|
Lacour et al. [68] |
Verbesserte Spine-Bildung durch EGb761® nach vestibulärer Schädigung |
|
Synapto(neurito)genese |
|
|
Xu et al. [137] |
EGb761® verbessert reduzierte CREB- und BDNF-Konzentrationen in Neuroblastomzellen nach Aβ-Behandlung. |
|
Tchantchou et al. [118] |
Erhöhtes Dendritenwachstum und erhöhte Konzentrationen von CREB und BDNF in hippocampalen Neuronen nach EGb761® |
|
Garcia-Alloza et al. [34] |
Verbessertes Neuritenwachstum durch EGb761® in einem AD-Maus-Modell |
|
Wang et al. [129] |
Verbessertes axonales Wachstum von retinalen Ganglienzellen durch Ginkgolid B |
|
Müller et al. [82] |
Verbesserte Neuritogenese von PC12-Zellen durch EGb761® und die drei Inhaltsstofffraktionen (Ginkgolide, Flavonoide, Bilobalid) |
|
Neurogenese |
|
|
Yoo et al. [138] |
Vermehrte Neurogenese im Hippocampus von adulten Mäusen nach 28-tägiger Behandlung mit Ginkgo-Extrakt (100 mg/kg) |
|
Tchantchou et al. [117] |
Vermehrte Neurogenese in einem AD-Maus-Modell nach 28-tägiger Behandlung mit EGb761® |
|
Tchantchou et al. [118] |
Verbesserte neuronale Proliferation nach subchronischer Behandlung mit EGb761® |
|
Nada et al. [83] |
Verbesserte Neurogenese nach experimentellem Hirninfarkt in der Maus |
|
Wang et al. [128] |
Verbesserte Neurogenese nach experimentellem Hirninfarkt in der Ratte durch Ginkgo-Extrakt |
AD: Alzheimer-Krankheit, BDNF: Brain-derived neurotrophic factor; CREB: Cyclic-AMP response element binding protein
Verbesserung der Kognition bei (noch) nicht dementen Patienten
Wie bereits erwähnt, sind kognitive Störungen mit zunehmendem Alter Teil eines Kontinuums, wo am Anfang leichtere kognitive, häufig auch stark subjektiv empfundene Beschwerden stehen, die sich über die Jahrzehnte bis zum Vollbild der Demenz mit ausgeprägten Defiziten entwickeln können. Über das ganze Kontinuum spielen mitochondriale Beeinträchtigungen und damit verbundene Störungen der neuronalen Funktion eine wesentliche Rolle. Die Frage, inwieweit EGb761® daher auch im Frühstadium, also zum Zeitpunkt noch nicht Diagnose-relevanter kognitiver Beeinträchtigungen, bei älteren Menschen positive Effekte zeigt, ist damit nicht nur für die Bestätigung des Konzepts wichtig, sondern spielt natürlich auch eine essenzielle Rolle bei der therapeutischen Anwendung von EGb761®, das ja als apothekenpflichtiges, aber nicht verschreibungspflichtiges Produkt eine breite Anwendung auch in diesem prädiagnostischen Bereich spielt. Die betroffenen Patienten sind natürlich daran interessiert, ihre leichten kognitiven Störungsbilder zu verbessern und gegebenenfalls auch eine Prävention dahingehend zu erreichen, dass der Übergang von leichteren kognitiven Einschränkungen in die weiteren Stadien dieses Kontinuums sich verlangsamt, vielleicht auch aufgehoben wird. Damit ist die klinische Frage, ob EGb761® auch positive Effekte bei diesen (noch) nicht kognitiv beeinträchtigten Patienten zeigt, von essenzieller Bedeutung für die Anwendung dieses Präparats.
Die Gruppen, die hier im Hinblick auf positive Effekte von EGb761® untersucht wurden, sind
- mittelalte, nicht kognitiv beeinträchtigte Probanden
- ältere Patienten mit sehr leichter oder milder kognitiver Beeinträchtigung (MCI; mild cognitive impairment) als wichtiger Vorstufe für eine spätere Alzheimer-Erkrankung im Hinblick auf eine noch mögliche kognitive Verbesserung und
- ältere nicht oder nur leicht kognitiv beeinträchtigte Menschen im Hinblick auf mögliche präventive Effekte für eine Verschlechterung der kognitiven Leistungsfähigkeit oder auf den Übergang zur MCI oder einer Alzheimer-Demenz.
Mittelalte, nicht kognitiv beeinträchtigte Probanden
In einer ersten Pilotstudie fanden Mix and Crews [75] positive Effekte von EGb761® (180 mg täglich über 6 Wochen) auf verschiedene Parameter von Kognition und Gedächtnis. Die Befunde konnten die Autoren in einer zweiten, größeren Untersuchung mit ähnlichem Design bestätigen [76]. Das mittlere Alter der Probanden betrug etwa 67 Jahre. Eine andere, auch über sechs Wochen gehende, allerdings nur 120 mg pro Tag einsetzende Studie fand dagegen keine positiven Effekte auf kognitive Parameter [109]. Neben der niedrigen Dosis wird die Studie besonders auch wegen der unsicheren Verblindung kritisiert. Eine neuere Untersuchung [61], ebenfalls über sechs Wochen, aber mit 240 mg EGb761® täglich an etwas jüngeren (etwa 55 Jahre) Probanden fand dagegen deutlichere Verbesserungen, besonders in einigen kognitiven Bereichen mit komplexen Aufgaben. Generell sind aber die Kognitions-verbessernden Effekte von EGb761® bei gesunden Probanden nicht sehr ausgeprägt, in Analogie zur Wirkung bei jungen Tieren.
Ältere Patienten mit vMCI oder MCI
Bei Patienten mit sehr leichten kognitiven Beeinträchtigungen (very mild cognitive impairment, vMCI) fanden Grass-Kapanke et al. [38] über verschiedene Bereiche des Kognitionsspektrums mäßige, aber signifikante Verbesserungen mit 240 mg EGb761® täglich über 12 Wochen. In einer weiteren Studie [36] an Patienten mit MCI mit neuropsychiatrischen Symptomen [7], wurden ebenfalls 240 mg EGb761® täglich über 24 Wochen gegeben. Auch hier wurden positive Effekte in verschiedenen kognitiven Domänen gesehen, darüber hinaus kam es zu einer Abnahme der neuropsychiatrischen Symptomatik.
Ältere nicht oder nur leicht kognitiv beeinträchtige Menschen
Aufgrund des heute akzeptierten Kontinuums von Hirnleistungsstörungen von der leichten kognitiven Störung hin bis zur klinisch manifesten Alzheimer-Demenz ist die Frage, ob man mit EGb761® im Frühstadium dahingehend eingreifen kann, dass die Progredienz verlangsamt wird, von großer Bedeutung. Zwei epidemiologische Studien [5, 6] geben erste Hinweise, dass die Einnahme von EGb761® über Jahre das Risiko, an einer Demenz zu erkranken, reduziert. Bedingt durch das kritisch zu sehende Design, besonders der neueren Studie, ergibt sich hier allerdings nur ein Hinweis, kein Beleg. Diesen hat man sich von zwei großen, über Jahre gehenden Studien erhofft, der in den USA durchgeführten GEM-Studie (Ginkgo evaluation of memory) und der in Frankreich durchgeführten GuidAge-Studie [132].
In der GEM-Studie wurden jeweils etwa 1500 sehr gesunde ältere Probanden (rund 80 Jahre) im Mittel mit 240 mg EGb761® pro Tag oder mit Placebo behandelt. Untersucht wurde das Risiko, an Alzheimer zu erkranken, oder die Abnahme der kognitiven Leistungsfähigkeit [24, 107]. Beide Endpunkte wurden von EGb761® nicht verändert. Obwohl diese Studie auf den ersten Blick beeindruckt, gibt es inzwischen erhebliche Zweifel an der Aussagekraft. Zum einen waren die relativ alten Teilnehmer trotz des hohen Alters kognitiv erstaunlich gesund und hatten auch in der Placebo-Gruppe nur minimale Veränderungen über die Zeit, was es einer pharmakologischen Intervention schwer macht, hier einen verlangsamenden Effekt zu zeigen. Zum anderen war die Adhärenz zur Medikation sehr schlecht; gemäß Erfassung der Tablettenblister lag sie bei nur 60%, war dann aber wahrscheinlich in Wirklichkeit noch geringer. Damit sind die Aussagen eher fraglich, da auch keine Korrekturen durchgeführt wurden.
Dass die Adhärenz tatsächlich relevant ist, zeigt die kleinere, aber im Detail sehr ähnliche Studie von Dodge et al. [28], die als Machbarkeitsstudie für die GEM gedacht war. Auch hier war in der Primärauswertung Ginkgo nicht wirksam, allerdings wurde eine signifikante Verlangsamung der Symptomatik über die Zeit (etwa 4 Jahre) unter Ginkgo gesehen, wenn die Auswertung für die Adhärenz, die hier mit etwa 75% sogar besser war, stratifiziert wurde. Eine solche eigentlich notwendige Stratifizierung wurde in der GEM-Studie nicht berichtet, was absolut unverständlich bleibt.
In der GuidAge-Untersuchung wurden etwa 2500 Patienten mit leichten kognitiven Beschwerden eingeschlossen und bei einer Tagesdosis von 240 mg über mehrere Jahre gegen Placebo beobachtet [120]. Während Ginkgo nach der ursprünglichen Auswertung keine gesicherten Effekte hatte, ergab sich nach einer Anpassung der Statistik an den tatsächlichen Verlauf (z.B. keine gleichmäßige Konversion in eine Demenz über den Verlauf) eine signifikante Reduktion des Demenzrisikos unter EGb761® für Patienten, die EGb761® mindestens vier Jahre eingenommen hatten (15 von 948 EGb761®; 28 von 963 Placebo). Warum diese Auswertung, obwohl sie im Studienplan als Sekundäranalyse aufgeführt war, in der Publikation fast versteckt wurde [120], bleibt ebenfalls offen.
Damit widerlegen beide Studien nicht eine präventive Wirkung von EGb761®, belegen sie aber auch nicht. Immerhin gibt die GuidAge-Studie ein Signal, wobei die lange benötigte Therapiezeit durchaus im Einklang zu den beiden bereits erwähnten epidemiologischen Studien steht.
Wirksamkeit bei Demenz
Obwohl EGb761 seit Jahrzehnten in der Behandlung von neurodegenerativen Demenzen erfolgreich eingesetzt wird, war die Bewertung in der Fachwelt für viele Jahre immer sehr zurückhaltend, zum Teil sogar negativ, wie beispielsweise in den Demenzleitlinien in Deutschland. Grund dafür war zum einen eine deutliche Heterogenität besonders bei älteren klinischen Studien mit zum Teil dann auch häufig überbewerteten negativen Daten, wie die amerikanische GEM-Studie mit erheblichen methodischen Mängeln, zum anderen aber sicher auch eine eher auf emotionaler Seite zu sehende negative Grundeinstellung vieler Kollegen gegenüber einem pflanzlichen Arzneimittel. Erst in den letzten Jahren, auch durch die konsequente Vorlage von immer neuen, modernen klinischen Standards entsprechenden Studien, hat sich die Einschätzung von EGb761® in der Behandlung von Demenzen in Leitlinien, Übersichtsarbeiten und Metaanalysen geändert (Tab. 4). Eine wichtige positive Bewertung wurde in Deutschland durch das IQWiG vorgelegt [55], dem man sicher keine emotionale Nähe zu pflanzlichen Arzneimitteln nachsagen kann. Ein weiterer Meilenstein war die positive, praktisch gleichwertige Einschätzung von EGb761® im Vergleich zu anderen Dementiva in den Leitlinien der Weltgesellschaften für Biologische Psychiatrie [50, 51]. Selbst die Empfehlungen für die neuen deutschen Leitlinien gehen von einer vorsichtig positiven Einschätzung von EGb761® in der Therapie der Demenz aus (S3-Leitlinie Demenzen, veröffentlicht Anfang 2016; Ia-Evidenz, Empfehlung „kann erwogen werden“).
Tab. 4. Die Akzeptanz von EGb761® als Antidementivum in offiziellen Leitlinien hat in den letzten 10 Jahren substanziell zugenommen
|
2015 |
Europäische Zulassungsbehörde bestätigt Wirksamkeit |
|
2014/16 |
Positive Bewertung in der S3-Leitlinie Demenzen |
|
2011 |
Empfehlung gleichwertig zu Acetylcholinesterase-(AChE-)Hemmern durch Weltverband für Biologische Psychiatrie |
|
2011 |
GKV-Therapiestandard Tagesdosis 240 mg |
|
2008 |
Positive Nutzenbewertung für EGb761® durch das IQWiG |
|
2000 |
Antidementivum nach ATC-Klassifikation, WHO |
Dieser Gesinnungswandel hat sicher viele Ursachen, im einfachsten Fall die in den letzten Jahren immer wieder vorgelegten positiven klinischen Daten, die sich dann auch in einer ganzen Reihe aktuellerer internationaler Metaanalysen mit positiver Bewertung niedergeschlagen haben [16, 35, 116, 122, 126, 130]. Sicher hat auch das Versagen der Amyloid-gerichteten Therapien dazu beigetragen, sich etwas stärker mit den bereits vorhandenen Therapien zu beschäftigen. Im Rahmen dieser Rückbesinnung ist man daher bereit, EGb761® einen adäquaten Platz in der Therapie der Alzheimer-Demenz neben den Acetylcholinesterasehemmern und Memantin zuzusprechen [51].
Es würde den Rahmen dieser Übersicht sprengen, auf die klinische Datenlage noch einmal detailliert einzugehen. Hier sei auf die bereits erwähnten neueren Metaanalysen zu EGb761® in der Behandlung der Alzheimer-Demenz hingewiesen, die alle von einer Wirksamkeit ausgehen. Anhand einiger repräsentativer Daten soll nur gezeigt werden, dass sich die Wirksamkeit sowohl auf kognitive Domänen der Alzheimer-Erkrankung wie auch auf die Alltagskompetenz niederschlägt.
Während verschiedene noch ältere Studien zum Teil methodisch anfechtbar waren, konnten die heute etwa 20 Jahre alten, damals aber nach neuesten Kriterien geplanten Studien von Kanowski et al. [58] und Le Bars et al. [69] klare kognitive Verbesserungen unter EGb761® zweifelsfrei zeigen (Abb. 6). Eine einige Jahre später durchgeführte Studie fand dagegen, unter ähnlichen Bedingungen durchgeführt, keine signifikante kognitive Verbesserung unter EGb761® im Vergleich zu Placebo [103]. Diese wurde allerdings bei einer Subgruppe der Patienten (etwa 30%) gefunden, die auch neuropsychiatrische Symptome (NPS) aufwiesen. Da NPS typisch für AD-Patienten sind (Abb. 7), wurden in der Folge vier Studien an Demenz-Patienten mit NPS durchgeführt (Tab. 5), wo deutliche Effekte auf Kognition und NPS gesehen wurden.
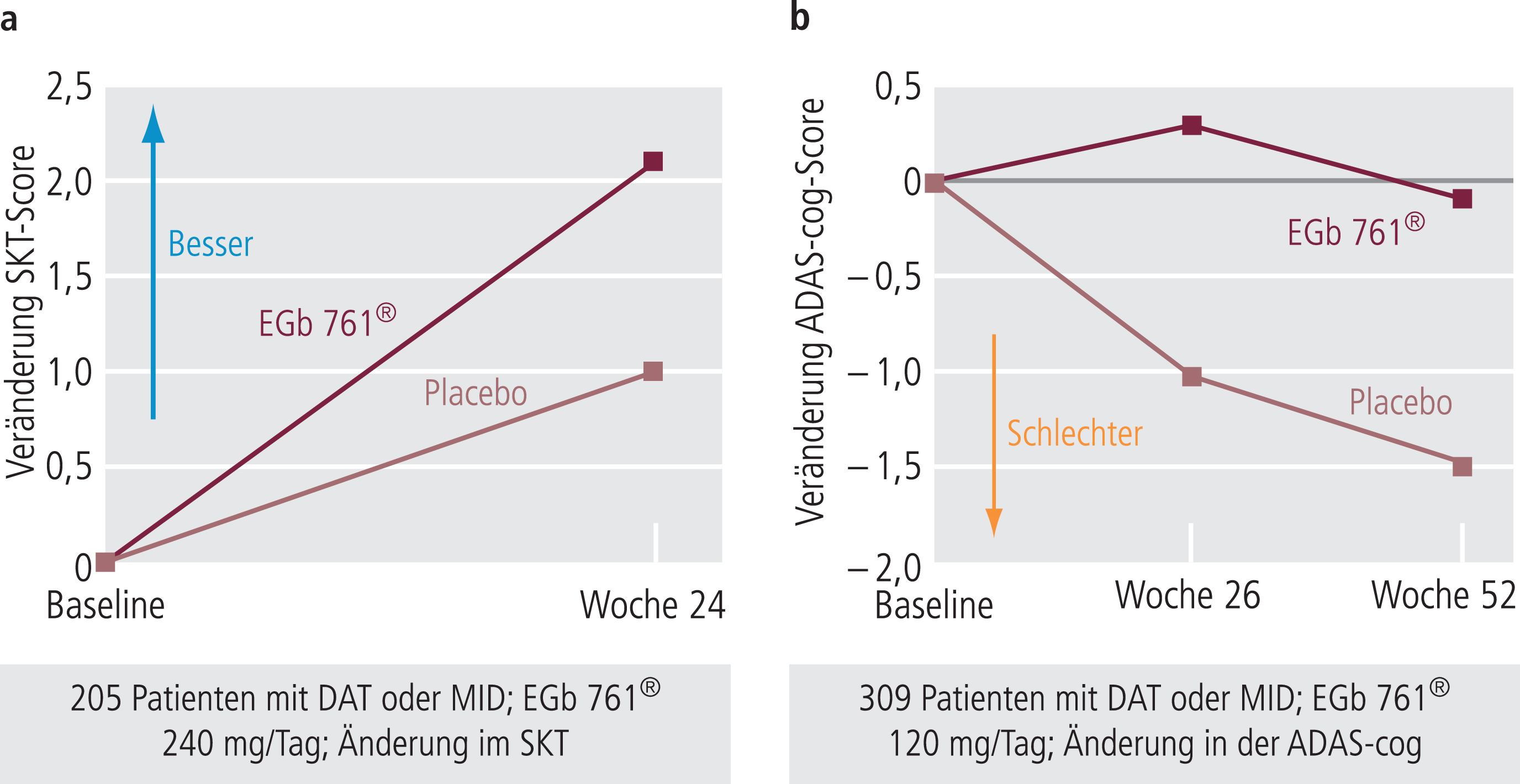
Abb. 6. Effekte von EGb761® in zwei großen Placebo-kontrollierten Doppelblindstudien in Deutschland bzw. den USA auf kognitive Störungen dementer Patienten unter Verwendung der SKT- bzw. ADAS-Cog-Skala [58, 69]. DAT: Demenz vom Alzheimer-Typ; MID: Multiinfarkt-Demenz; SKT: Syndrom-Kurztest
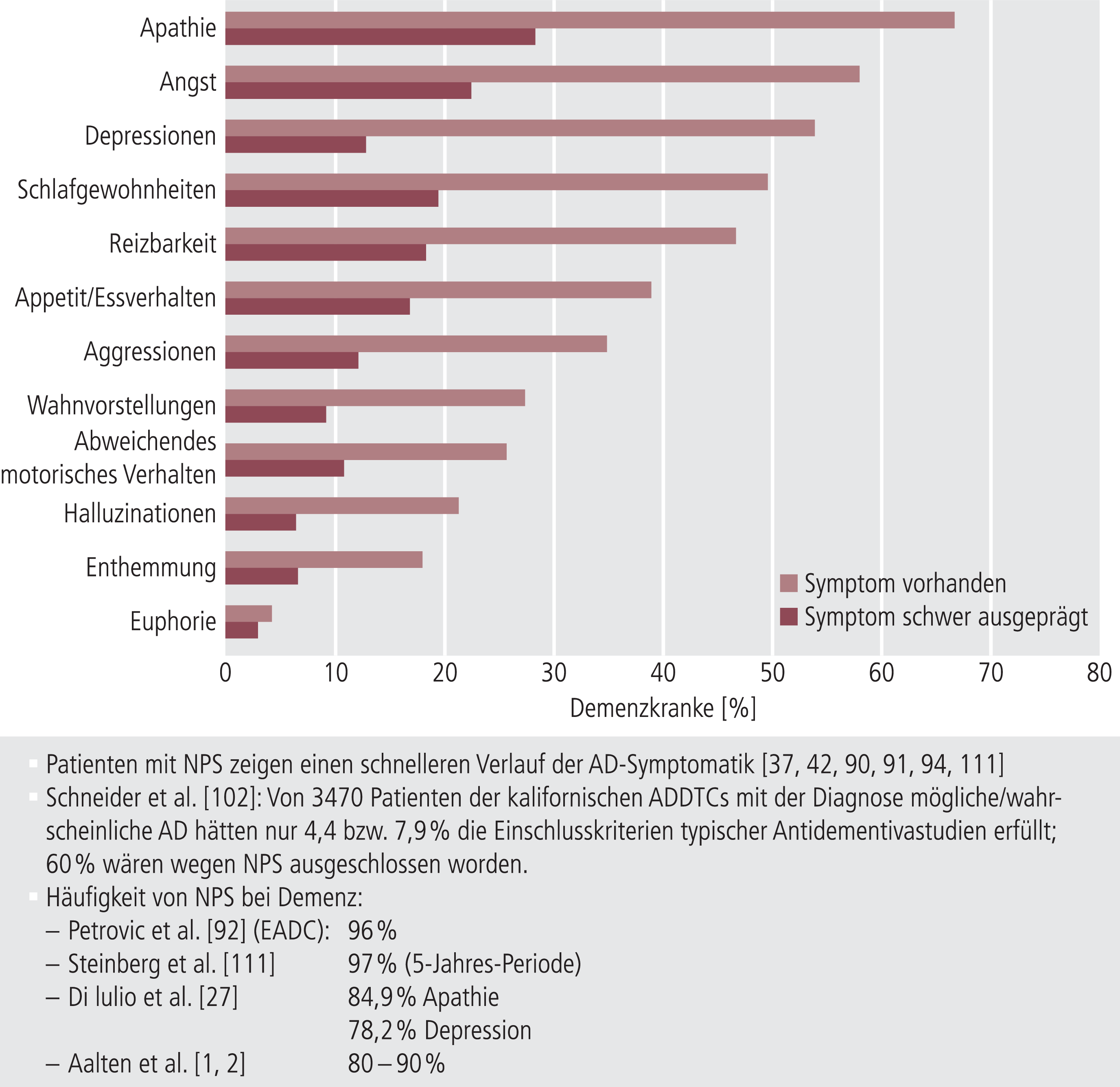
Abb. 7. Neuropsychiatrische Symptome (NPS) sind bei Demenzen (Alzheimer [AD] oder vaskulär) sehr häufig vorhanden und gut belegte Prädiktoren für einen ungünstigen Verlauf. ADDTCs: Alzheimer’s Disease Diagnostic and Treatment Centers; EADC: European Alzheimer’s Disease Consortium
Tab. 5. Neuere Therapie-Studien mit EGb761® bei Demenz bei Patienten mit neuropsychiatrischen Symptomen (NPS) als „enriched“ Studienpopulation. Kognitive Parameter wurden in allen vier Studien mit der SKT erfasst.
|
Vier randomisierte, Placebo-kontrollierte Studien:
|
|
Patienten: Indikation Alzheimer-Demenz oder vaskuläre Demenz |
|
Behandlung: 240 mg/Tag EGb 761® über 22 bzw. 24 Wochen |
|
Ausgewertete Zielgrößen:
|
Die Daten von zwei im Design identischen Studien sind in Abbildung 8 dargestellt. Auch eine Studie an MCI-Patienten hat mit diesem „enriched design“ gute Ergebnisse gebracht [36].
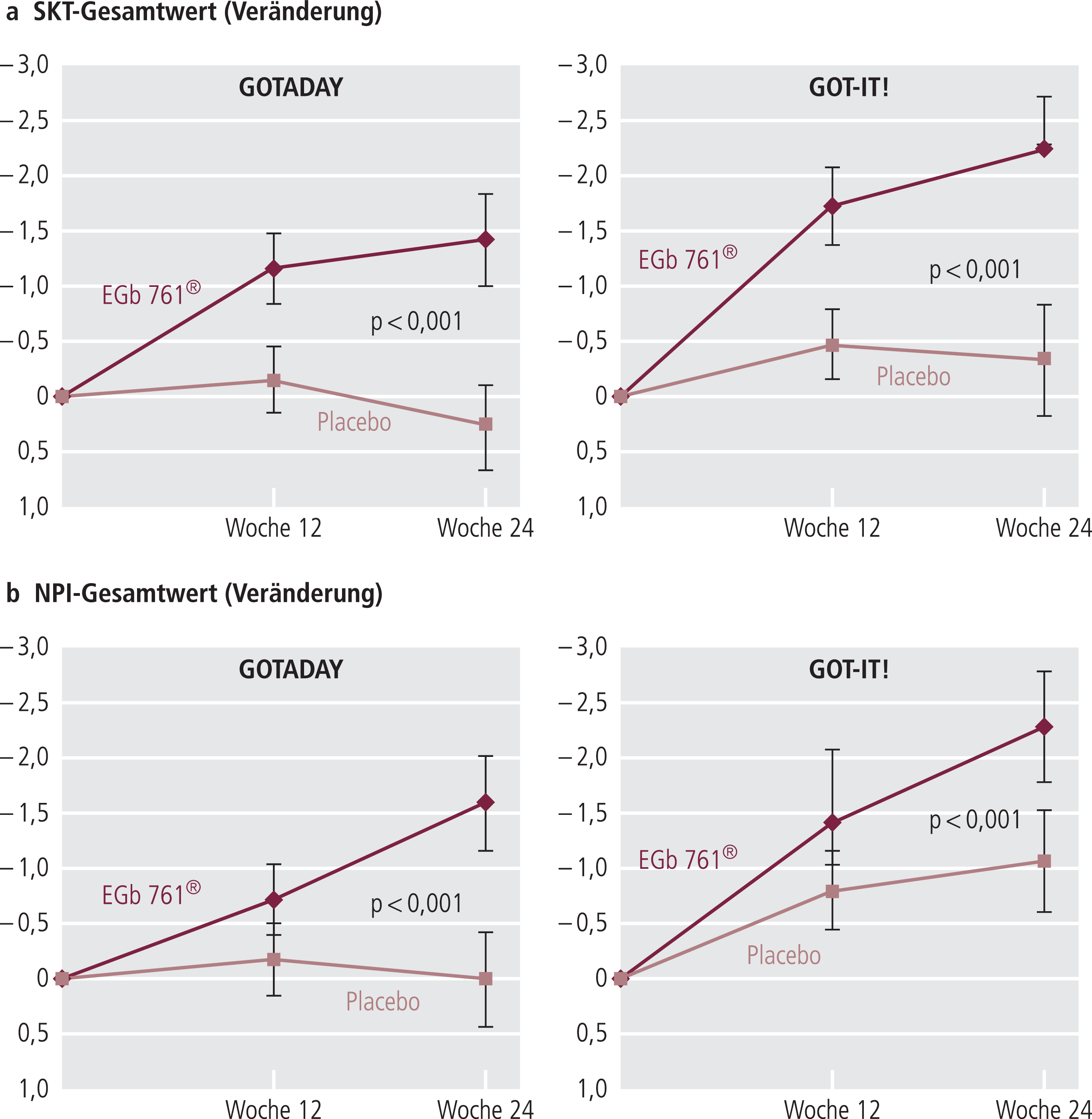
Abb. 8. Konsistente Verbesserung im SKT und NPI in zwei im Design identischen Studien (GOTADAY, GOT-IT!) mit EGb761® bei Demenz-Patienten [43, 49]
EGb761® zeigt darüber hinaus sehr deutliche Effekte besonders auf den affektiven Teil der neuropsychiatrischen Begleitsymptomatik [60, 105].
Verschiedene Sekundäranalysen zeigen, dass die beobachtete Wirksamkeit von EGb761® in der Responderrate, der NNT-Auswertung (Number needed to treat) und der Verzögerung der Symptomprogression im Rahmen der Werte von den anderen Antidementiva (Acetylcholinesterasehemmer, Memantin) liegt (Abb. 9) [62, 63]. Dies wird auch durch eine Vergleichsstudie bestätigt [137]. Als großer Vorteil ist die unbestritten gute Verträglichkeit zu bewerten, ebenso die ähnlich gute Wirksamkeit bei Alzheimer-Demenz und vaskulärer Demenz (siehe die vergleichende Betrachtung von Napryeyenko et al. [85]).
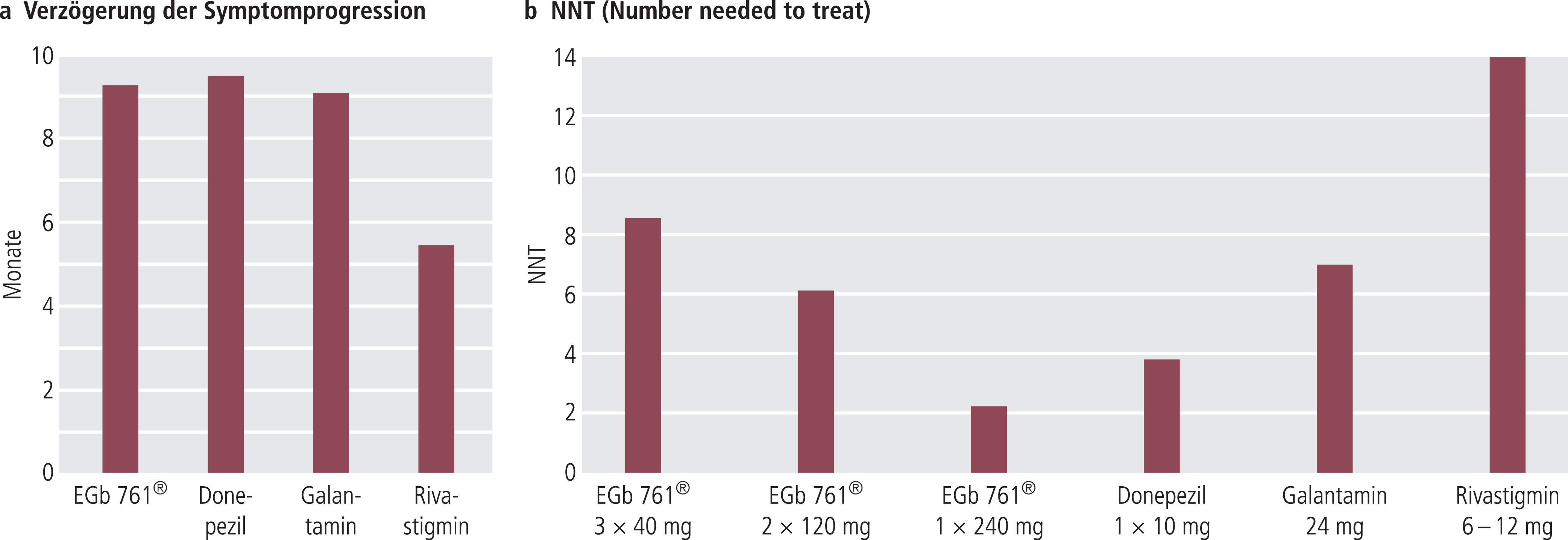
Abb. 9. Die Wirksamkeit von EGb761® im Vergleich zu Acetylcholinesterasehemmern; a) Verzögerung der Symptomprogression bei Alzheimer-Demenz unter EGb761® und Cholinesterasehemmern (EGb761® 120 mg/Tag, Donepezil 10 mg/Tag, Galantamin 24 mg/Tag, Rivastigmin 6–12 mg/Tag [62, 63]; b) Numbers needed to treat (NNT) für EGb761® und die Acetylcholinesterasehemmer Donepezil, Galantamin, Rivastigmin. Die Daten wurden aus ITT-Analysen zusammengestellt [62, 63]
Schlussfolgerung und Zusammenfassung
Der standardisierte Ginkgo-Extrakt EGb761® (Tebonin®) wurde 1965 zur Behandlung von peripheren und besonders auch von zentralen Durchblutungsstörungen eingesetzt. Schon bald zeigte sich die Wirksamkeit bei altersbedingten Hirnleistungsstörungen, die durch immer neue den jeweiligen Diagnose-Standards angepassten klinischen Studien bis heute bestätigt wurde. Dies hat sich ganz aktuell in verschiedenen Leitlinien niedergeschlagen, die EGb761® als in der klinischen Wirksamkeit bei Alzheimer und vaskulären Demenzen belegtes Arzneimittel aufführen. Parallel zu den aktuellen Studien zur klinischen Wirksamkeit konnte in experimentellen Studien gezeigt werden, dass EGb761® hauptsächlich über eine Verbesserung der mitochondrialen Funktion wirkt. Die damit verbesserte Versorgung der zentralen Neuronen mit energiereichen Phosphaten führt zu einer Reduktion typischer neurochemischer und histologischer Defizite des alten Gehirns bezüglich Neurotransmission und Neuroplastizität. Da diese Störungen nach dem Debakel mit der Amyloid-Hypothese heute als wesentliche Ursache von Alzheimer-Demenz und vaskulärer Demenz gelten, kann man heute, 50 Jahre nach Einführung von EGb761®, davon ausgehen, dass der Pflanzenextrakt spezifisch wichtige Aspekte der Pathophysiologie der Demenz verbessert.
Interessenkonflikterklärung
WEM: Vortrags- und Beraterhonorare von Dr. Willmar Schwabe GmbH & Co KG (Karlsruhe) und Lundbeck (Hamburg) sowie Forschungsunterstützung von Dr. Willmar Schwabe GmbH & Co KG (Karlsruhe).
AE: Vortragshonorare im Rahmen wissenschaftlicher Symposium (Satellitensymposien bei Fachtagungen/-Konferenzen) von der Schwabe Pharma AG, Küssnacht am Rigi, Schweiz, und der Dr. Willmar Schwabe GmbH & Co. KG, Karlsruhe, Deutschland.
GPE: Es bestehen keine Interessenkonflikte.
HF: Auftragsforschung für Firma Schwabe
KF: Forschungsunterstützung von Dr.Willmar Schwabe GmbH&Co.KG, Karlsruhe.
RH: Wissenschaftlicher Mitarbeit der Dr.Willmar Schwabe GmbH&Co.KG, Karlsruhe.
RI: Forschungsbeihilfen, Honorare für Beratung oder Teilnahme an einem Expertenbeirat, Vortragshonorare oder Autorenhonorare von APK, Austroplant, Beltz Test, BMG, BMFSFJ, BOD, Caritas Siegen, DKV, European Commission, Friedrichverlag, Hogrefe, LVR, Schwabe, Servier, Springer-Verlag, Thieme, Urban & Vogel, WDR.
SK: Forschungsstützung oder Honorare von Angelini, AOP Orphan Pharmaceuticals AG, AstraZeneca, Krka-Pharma, Eli Lilly, Janssen, Lundbeck, Neuraxpharm, Pfizer, Pierre Fabre, Schwabe und Servier.
HJM: Honorare für Vortrags- und Beratertätigkeiten oder Forschungsgelder von Astra-Zeneca, Eli Lilly, Janssen, Lundbeck, Pfizer, Schwabe, Servier, Otsuka und Takeda. HJM war Präsident oder Vorstandsmitglied der CINP, ECNP, WFSBP und EPA und Vorsitzender der WPA-Sektion zur Pharmakopsychiatrie.
Literatur
1. Aalten P, Verhey FR, Boziki M, Bullock R, et al. Neuropsychiatric syndromes in dementia. Results from the European Alzheimer Disease Consortium: Part I. Dement Geriatr Cogn Disord 2007;24:457–63.
2. Aalten P, Verhey FR, Boziki M, Brugnolo A, et al. Consistency of neuropsychiatric syndromes across dementias: results from the European Alzheimer Disease Consortium. Part II. Dement Geriatr Cogn Disord 2008;25:1–8.
3. Abdel-Kader R, Hauptmann S, Keil U, Scherping I, et al. Stabilization of mitochondrial function by Ginkgo biloba extract (EGb 761). Pharmacol Res 2007;56:493–502.
4. Ahlemeyer B, Möwes A, Krieglstein J. Inhibition of serum deprivation- and staurosporine-induced neuronal apoptosis by Ginkgo biloba extract and some of its constituents. Eur J Pharmacol 1999;367:423–30.
5. Amieva H, Meillon C, Helmer C, Barberger-Gateau P, et al. Ginkgo biloba extract and long-term cognitive decline: a 20-year follow-up population-based study. PLoS One 2013;8:e52755.
6. Andrieu S, Gillette S, Amouyal K, Nourhashemi F, et al.; EPIDOS study. Association of Alzheimer’s disease onset with ginkgo biloba and other symptomatic cognitive treatments in a population of women aged 75 years and older from the EPIDOS study. J Gerontol A Biol Sci Med Sci 2003;58:372–7.
7. Apostolova LG, Cummings JL. Neuropsychiatric manifestations in mild cognitive impairment: a systematic review of the literature. Dement Geriatr Cogn Disord 2008;25:115–26.
8. Baki L, Neve RL, Shao Z, Shioi J, et al. Wild-type but not FAD mutant presenilin-1 prevents neuronal degeneration by promoting phosphatidylinositol 3-kinase neuroprotective signalling. J Neurosci 2008;28:483–90.
9. Baliutyte G, Trumbeckaite S, Baniene R, Borutaite V, et al. Effects of standardized extract of Ginkgo biloba leaves EGb761 on mitochondrial functions: mechanism(s) of action and dependence on the source of mitochondria and respiratory substrate. J Bioenerg Biomembr 2014;46:493–501.
10. Barthet G, Dunys J, Shao Z, Xuan Z, et al. Presenilin mediates neuroprotective functions of ephrin B and brain-derived neurotrophic factor and regulates ligand-induced internalization and metabolism of EphB2 and TrkB receptors. Neurobiol Aging 2013;34:499–510.
11. Bastianetto S1, Ramassamy C, Doré S, Christen Y, et al. The Ginkgo biloba extract (EGb 761) protects hippocampal neurons against cell death induced by beta-amyloid. Eur J Neurosci 2000;12:1882–90.
12. Belviranlı M, Okudan N. The effects of Ginkgo biloba extract on cognitive functions in aged female rats: The role of oxidative stress and brain-derived neurotrophic factor. Behav Brain Res 2015;278:453–61.
13. Berger DM. The diagnosis of organic brain syndrome. Can Fam Physion 1977;23:344–7.
14. Biber A. Pharmacokinetics of Ginkgo biloba extracts. Pharmacopsychiatry 2003;36(Suppl 1): S32–7.
15. Blecharz-Klin K, Piechal A, Joniec I, Pyrzanowska J, et al. Pharmacological and biochemical effects of Ginkgo biloba extract on learning, memory consolidation and motor activity in old rats. Acta Neurobiol Exp (Wars) 2009;69:217–31.
16. Brondino N, De Silvestri A, Re S, Lanati N, et al. A systematic review and meta-analysis of Ginkgo biloba in neuropsychiatric disorders: From ancient tradition to modern-day medicine. Evid Based Complement Alternat Med 2013;2013:915691.
17. Bruban J, Voloudakis G, Huang Q, Kajiware Y, et al. Presenilin 1 is necessary for neuronal, but not glial, EGFR expression and neuroprotection via γ-secretase-independent transcriptional mechanisms. FASEB J 2015;29:3702–12.
18. Caesar W. [Of Ginkgo EGb on GBL – a long path to rational phytopharmacy]. Pharm Unserer Zeit 2009;38:400–5.
19. Chen HK, Ji ZS, Dodson SE, Miranda RD, et al. Apolipoprotein E4 domain interaction mediates detrimental effects on mitochondria and is a potential therapeutic target for Alzheimer disease. J Biol Chem 2011;286:5215–21.
20. Colciaghi F, Borroni B, Zimmermann M, Bellone C, et al. Amyloid precursor protein metabolism is regulated toward alpha-secretase pathway by Ginkgo biloba extracts. Neurobiol Dis 2004;16:454–60.
21. Commenges D, Scotet V, Renaud S, Jacqmin-Gadda H, et al. Intake of flavonoids and risk of dementia. Eur J Epidemiol 2000;16:357–63.
22. DeFeudis FV. A brief history of EGb761 and its therapeutic uses. Pharmacopsychiatry 2003;36(Suppl 1):S2–7.
23. DeFeudis FV. Ginkgo biloba extract (EGb761). From chemistry to the clinic. Wiesbaden: Ullstein Medical, 1998.
24. DeKosky ST, Williamson JD, Fitzpatrick AL, Kronmal RA, et al.; Ginkgo Evaluation of Memory (GEM) Study Investigators. Ginkgo biloba for prevention of dementia: a randomized controlled trial. JAMA 2008;300:2253–62.
25. Devore EE, Grodstein F, van Rooij FJ, Hofman A, et al. Dietary antioxidants and long-term risk of dementia. Arch Neurol 2010;67:819–25.
26. Diamond BJ, Shiflett SC, Feiwel N, Matheis RJ, et al. Ginkgo biloba extract: mechanisms and clinical indications. Arch Phys Med Rehabil 2000;81:668–78.
27. Di Lulio F, Palmer K, Blundo C, Casini AR, et al. Occurrence of neuropsychiatric symptoms and psychiatric disorders in mild Alzheimer’s disease and mild cognitive impairment subtypes. Int Psychogeriatr 2010;22:629–40.
28. Dodge HH, Zitzelberger T, Oken BS, Howieson D, et al. A randomized placebo-controlled trial of Ginkgo biloba for the prevention of cognitive decline. Neurology 2008;70:1809–17.
29. Eckert A, Keil U, Kressmann S, Schindowski K, et al. Effects of EGb761 Ginkgo biloba extract on mitochondrial function and oxidative stress. Pharmacopsychiatry 2003;36(Suppl 1):S15–23.
30. Eckert A, Keil U, Scherping I, Hauptmann S, et al. Stabilization of mitochondrial membrane potential and improvement of neuronal energy metabolism by Ginkgo biloba extract EGb761. Ann N Y Acad Sci 2005;1056:474–85.
31. Engelhart MJ, Geerlings MI, Ruitenberg A, van Swieten JC, et al. Dietary intake of antioxidants and risk of Alzheimer disease. JAMA 2002;287:3223–9.
32. Epstein LJ, Simon A. Organic brain syndrome in the elderly. Geriatrics 1967;22:145–50.
33. Fehske CJ, Leuner K, Müller WE. Ginkgo biloba extract (EGb761) influences monoaminergic neurotransmission via inhibition of NE uptake, but not MAO activity after chronic treatment. Pharmacol Res 2009;60:68–73.
34. Garcia-Alloza M, Borrelli LA, Hyman BT, Bacskai BJ. Antioxidants have a rapid and long-lasting effect on neuritic abnormalities in APP:PS1 mice. Neurobiol Aging 2010;31:2058–68.
35. Gauthier S, Schlaefke S. Efficacy and tolerability of Ginkgo biloba extract EGb761® in dementia: a systematic review and meta-analysis of randomized placebo-controlled trials. Clin Interv Aging 2014;9:2065–77.
36. Gavrilova SI, Preuss UW, Wong JW, Hoerr R, et al.; GIMCIPlus Study Group. Efficacy and safety of Ginkgo biloba extract EGb761 in mild cognitive impairment with neuropsychiatric symptoms: a randomized, placebo-controlled, double-blind, multi-center trial. Int J Geriatr Psychiatry 2014;29:1087–95.
37. Geda YE, Roberts RO, Mielke MM, Knopman DS, et al. Baseline neuropsychiatric symptoms and the risk of incident mild cognitive impairment: a population-based study. Am J Psychiatry 2014;171:572–81.
38. Grass-Kapanke B, Busmane A, Lasmanis A, Hoerr R, et al. Effects of ginkgo biloba special extract EGb761 in very mild cognitive impairment (vMCI). Neuroscience & Medicine 2011;2;48–56.
39. Halama P, Bartsch G, Meng G. [Disorders of brain performance of vascular origin. Randomized double-blind study of the effectiveness of Gingko biloba extract]. Fortschr Med 1988;106:408–12.
40. Hamann KF. [Special ginkgo extract in cases of vertigo: a systematic review of randomised, double-blind, placebo controlled clinical examinations]. HNO 2007;55:258–63.
41. Heiß WD, Zeiler K. Medikamentöse Beeinflussung der Hirndurchblutung. Pharmakotherapie 1978;1:137–44.
42. Herrmann N, Harimoto T, Balshaw R, Lanctôt KL; Canadian Outcomes Study in Dementia (COSID) Investigators. Risk factors for progression of Alzheimer disease in a Canadian population: The Canadian Outcomes Study in Dementia (COSID). Can J Psychiatry 2015;60:189–99.
43. Herrschaft H, Nacu A, Likhachev S, Sholomov I, et al. Ginkgo biloba extract EGb761® in dementia with neuropsychiatric features: a randomised, placebo-controlled trial to confirm the efficacy and safety of a daily dose of 240 mg. J Psychiatr Res 2012;46:716–23.
44. Hoerr R, Zaudig M. A retrospective classification of diagnoses in terms of DSM-5 for patients included in randomized controlled trials of Ginkgo biloba extract EGb761®. Eur Arch Psychiatry 2016;266:249–59.
45. Hofferberth B. Simultanerfassung elektrophysiologischer, psychometrischer und rheologischer Parameter bei Patienten mit hirnorganischem Psychosyndrom und erhöhtem Gefäßrisiko – Eine Placebo-kontrollierte Doppelblindstudie mit Ginkgo-biloba-Extrakt EGb761®. In: Stodtmeister KR, Pillunat LE (eds.). Mikrozirkulation in Gehirn und Sinnesorganen. Stuttgart: Enke, 1991:64–74.
46. Hoffman SW, Stein DG. Extract of Ginkgo biloba (EGb761) improves behavioral performance and reduces histopathology after cortical contusion in the rat. Restor Neurol Neurosci 1997;11:1–12.
47. Hoyer S, Lannert H, Nöldner M, Chatterjee SS. Damaged neuronal energy metabolism and behavior are improved by Ginkgo biloba extract (EGb761). J Neural Transm 1999;106:1171–88.
48. Huang Y. Abeta-independent roles of apolipoprotein E4 in the pathogenesis of Alzheimer’s disease. Trends Mol Med 2010;16:287–94.
49. Ihl R, Bachinskaya N, Korczyn AD, Vakhapova V, et al; GOTADAY Study Group. Efficacy and safety of a once-daily formulation of Ginkgo biloba extract EGb761 in dementia with neuropsychiatric features: a randomized controlled trial. Int J Geriatr Psychiatry 2011;26:1186–94.
50. Ihl R, Bunevicius R, Frölich L, Winblad B, et al.; WFSBP Task Force on Mental Disorders in Primary Care; WFSBP Task Force on Dementia. World Federation of Societies of Biological Psychiatry guidelines for the pharmacological treatment of dementias in primary care. Int J Psychiatry Clin Pract 2015;19:2–7.
51. Ihl R, Frölich L, Winblad B, Schneider L, et al.; WFSBP Task Force on Treatment Guidelines for Alzheimer’s Disease and other Dementias. World Federation of Societies of Biological Psychiatry (WFSBP) guidelines for the biological treatment of Alzheimer’s disease and other dementias. World J Biol Psychiatry 2011;12:2–32.
52. Iqbal K, Liu F, Gong CX. Alzheimer disease therapeutics: focus on the disease and not just plaques and tangles. Biochem Pharmacol 2014;88:631–9.
53. Janssens D, Michiels C, Delaive E, Eliaers F, et al. Protection of hypoxia-induced ATP decrease in endothelial cells by ginkgo biloba extract and bilobalide. Biochem Pharmacol 1995;50:991–9.
54. Jack CR, Wiste HJ, Weigend SD, Rocca WA, et al. Age-specific population frequencies of cerebral β-amyloidosis and neurodegeneration among people with normal cognitive function aged 50–89 years: a cross-sectional study. Lancet Neurol 2014;13:997–1005.
55. Janssen IM, Sturtz S, Skipka G, Zentner A, et al. Ginkgo biloba in Alzheimer’s disease: a systematic review. Wien Med Wochenschr 2010;160:539–46.
56. Janssens D, Remacle J, Drieu K, Michiels C. Protection of mitochondrial respiration activity by bilobalide. Biochem Pharmacol 1999;58:109–19.
57. Kanowski S. Clinical and pathological aspects of chronic organic brain syndrome. Exp Brain Res 1982;(Suppl 5):223–34.
58. Kanowski S, Herrmann WM, Stephan K, Wierich W, et al. Proof of efficacy of the ginkgo biloba special extract EGb761 in outpatients suffering from mild to moderate primary degenerative dementia of the Alzheimer type or multi-infarct dementia. Pharmacopsychiatry 1996;29:47–56.
59. Karran E, Hardy J. A critique of the drug discovery and phase 3 clinical programs targeting the amyloid hypothesis for Alzheimer disease. Ann Neurol 2014;76:185–205.
60. Kaschel R. Ginkgo biloba: specificity of neuropsychological improvement – a selective review in search of differential effects. Hum Psychopharmacol 2009;24:345–70.
61. Kaschel R. Specific memory effects of Ginkgo biloba extract EGb761 in middle-aged healthy volunteers. Phytomedicine 2011;18:1202–7.
62. Kasper S, Schubert H. [Ginkgo biloba extract EGb761 in the treatment of dementia: evidence of efficacy and tolerability]. Fortschr Neurol Psychiatr 2009;77:494–506.
63. Kasper S, Winkler D, Kutzelnigg A. [Significance of Ginkgo biloba extract in dementia]. Pharm Unserer Zeit 2009;38:424–30.
64. Kellermann AJ, Kloft C. Is there a risk of bleeding associated with standardized Ginkgo biloba extract therapy? A systematic review and meta-analysis. Pharmacotherapy 2011;31:490–502.
65. Kehr J, Yoshitake S, Ijiri S, Koch E, et al. Ginkgo biloba leaf extract (EGb761®) and its specific acylated flavonol constituents increase dopamine and acetylcholine levels in the rat medial prefrontal cortex: possible implications for the cognitive enhancing properties of EGb761®. Int Psychogeriatr 2012;24(Suppl 1):S25–34.
66. Kloft C, Kellermann A. [Bleeding caused by Ginkgo biloba extract?]. Pharm Unserer Zeit 2009;38:440–6.
67. Koch E. Inhibition of platelet activating factor(PAF)-induced aggregation of human thrombocytes by ginkgolides: considerations on possible bleeding complications after oral intake of Ginkgo biloba extracts. Phytomedicine 2005;12:10–6.
68. Lacour M, Ez-Zaher L, Raymond J. Plasticity mechanisms in vestibular compensation in the cat are improved by an extract of Ginkgo biloba (EGb761). Pharmacol Biochem Behav 1991;40:367–79.
69. Le Bars PL, Katz MM, Berman N, Itil TM, et al. A placebo-controlled, double-blind, randomized trial of an extract of Ginkgo biloba for dementia. North American EGb Study Group. JAMA 1997;278:1327–32.
70. Leuner K, Hauptmann S, Abdel-Kader R, Scherping I, et al. Mitochondrial dysfunction: the first domino in brain aging and Alzheimer’s disease? Antioxid Redox Signal 2007;9:1659–75.
71. Leuner K, Müller WE, Reichert AS. From mitochondrial dysfunction to amyloid beta formation: novel insights into the pathogenesis of Alzheimer’s disease. Mol Neurobiol 2012;46:186–93.
72. Leuner K, Schütt T, Kurz C, Eckert SH, et al. Mitochondrion-derived reactive oxygen species lead to enhanced amyloid beta formation. Antioxid Redox Signal 2012;16:1421–33.
73. Lin CC, Cheng WL, Hsu SH, Chang CM. The effects of Ginkgo biloba extracts on the memory and motor functions of rats with chronic cerebral insufficiency. Neuropsychobiology 2003;47:47–51.
74. Liu X, Hao W, Qin Y, Decker Y, et al. Long-term treatment with Ginkgo biloba extract EGb761 improves symptoms and pathology in a transgenic mouse model of Alzheimer’s disease. Brain Behav Immun 2015;46:121–31.
75. Mix JA, Crews WD Jr. An examination of the efficacy of Ginkgo biloba extract EGb761 on the neuropsychologic functioning of cognitively intact older adults. J Altern Complement Med 2000;6:219–29.
76. Mix JA, Crews WD Jr. A double-blind, placebo-controlled, randomized trial of Ginkgo biloba extract EGb761 in a sample of cognitively intact older adults: neuropsychological findings. Hum Psychopharmacol 2002;17:267–77.
77. Montes P, Ruiz-Sanchez E, Rojas C, Rojas P. Ginkgo biloba extract 761: A review of basic studies and potential clinical use in psychiatric disorders. CNS Neurol Disord Drug Targets 2015;14:132–49.
78. Moreau P. [A new stimulating agent of cerebrovascular circulation]. Nouv Presse Med 1975;4:2401–2.
79. Müller WE, Juretzek W. Ginkgo biloba. In: Riederer P, Laux G, Pöldinger W (Hrsg.). Neuro-Psychopharmaka. Wien: Springer-Verlag, 1999.
80. Müller WE, Chatterjee SS. Cognitive and other behavioral effects of EGb761 in animal models. Pharmacopsychiatry 2003;36(Suppl 1): S24–31.
81. Müller WE, Heiser J, Leuner K. Effects of the standardized Ginkgo biloba extract EGb761® on neuroplasticity. Int Psychogeriatr 2012;24(Suppl 1):S21–4.
82. Müller WE, Eckert A, Kurz C, Eckert GP, et al. Mitochondrial dysfunction: common final pathway in brain aging and Alzheimer’s disease – therapeutic aspects. Mol Neurobiol 2010;41:159–71.
83. Nada SE, Tulsulkar J, Shah ZA. Heme oxygenase 1-mediated neurogenesis is enhanced by Ginkgo biloba (EGb761®) after permanent ischemic stroke in mice. Mol Neurobiol 2014;49:945–56.
84. Napryeyenko O, Borzenko I; GINDEM-NP Study Group. Ginkgo biloba special extract in dementia with neuropsychiatric features. A randomised, placebo-controlled, double-blind clinical trial. Arzneimittelforschung 2007;57:4–11.
85. Napryeyenko O, Sonnik G, Tartakovsky I. Efficacy and tolerability of Ginkgo biloba extract EGb761 by type of dementia: analyses of a randomised controlled trial. J Neurol Sci 2009;283:224–9.
86. Nikolova G, Yancheva S, Raychev I, Hoerr R, for the Plagin Study Group. Ginkgo biloba extract in dementia: A 22-week randomized, placebo-controlled, double-blind trial. Bulgarian Neurology 2013;14:139–43.
87. Oliveira DR, Sanada PF, Saragossa Filho AC, Innocenti LR, et al. Neuromodulatory property of standardized extract Ginkgo biloba L. (EGb761) on memory: behavioral and molecular evidence. Brain Res 2009;1269:68–89.
88. Paganelli RA, Benetoli A, Milani H. Sustained neuroprotection and facilitation of behavioral recovery by the Ginkgo biloba extract, EGb761, after transient forebrain ischemia in rats. Behav Brain Res 2006;174:70–7.
89. Pardon MC, Hanoun N, Perez-Diaz F, Joubert C, et al. Long-term treatment with the antioxidant drug EGb761 at senescence restored some neurobehavioral effects of chronic ultramild stress exposure seen in young mice. Neurobiol Aging 2004;25:1067–83.
90. Peters ME, Rosenberg PB, Steinberg M, Norton MC, et al.; Cache County Investigators. Neuropsychiatric symptoms as risk factors for progression from CIND to dementia: the Cache County Study. Am J Geriatr Psychiatry 2013;21:1116–24.
91. Peters ME, Schwartz S, Han D, Rabins PV, et al. Neuropsychiatric symptoms as predictors of progression to severe Alzheimer’s dementia and death: the Cache County Dementia Progression Study. Am J Psychiatry 2015;172:460–5.
92. Petrovic M, Hurt C, Collins D, Burns A, et al. Clustering of behavioural and psychological symptoms in dementia (BPSD): a European Alzheimer’s disease consortium (EADC) study. Acta Clin Belg 2007;62:426–32.
93. Porsolt RD, Martin P, Lenègre A, Fromage S, et al. Effects of an extract of Ginkgo Biloba (EGB 761) on “learned helplessness” and other models of stress in rodents. Pharmacol Biochem Behav 1990;36:963–71.
94. Rabins PV, Schwartz S, Black BS, Corcoran C, et al. Predictors of progression to severe Alzheimer’s disease in an incidence sample. Alzheimers Dement 2013;9:204–7.
95. Reddy PH. Mitochondrial dysfunction in aging and Alzheimer’s disease: strategies to protect neurons. Antioxid Redox Signal 2007;9:1647–58.
96. Rhein V, Giese M, Baysang G, Meier F, et al. Ginkgo biloba extract ameliorates oxidative phosphorylation performance and rescues abeta-induced failure. PLoS One 2010;5:e12359.
97. Sastre J, Lloret A, Borrás C, Pereda J, et al. Ginkgo biloba extract EGb761 protects against mitochondrial aging in the brain and in the liver. Cell Mol Biol (Noisy-le-grand) 2002;48:685–92.
98. Satvat E, Mallet PE. Chronic administration of a Ginkgo biloba leaf extract facilitates acquisition but not performance of a working memory task. Psychopharmacology (Berl) 2009;202:173–85.
99. Scheff SW, Price DA, Schmitt FA, DeKosky ST, et al. Synaptic alterations in CA1 in mild Alzheimer disease and mild cognitive impairment. Neurology 2007;68:1501–8.
100. Scheff SW, Price DA, Schmitt FA, Roberts KN, et al. Synapse stability in the precuneus early in the progression of Alzheimer’s disease. J Alzheimers Dis 2013;35:599–609.
101. Schindowski K, Leutner S, Kressmann S, Eckert A, et al. Age-related increase of oxidative stress-induced apoptosis in mice prevention by Ginkgo biloba extract (EGb761). J Neural Transm 2001;108:969–78.
102. Schneider LS, Olin JT, Lyness SA, Chui HC. Eligibility of Alzheimer’s disease clinic patients for clinical trials. J Am Geriatr Soc 1997;45:923–8.
103. Schneider LS, DeKosky ST, Farlow MR, Tariot PN, et al. A randomized, double-blind, placebo-controlled trial of two doses of Ginkgo biloba extract in dementia of the Alzheimer’s type. Curr Alzheimer Res 2005;2:541–51.
104. Schuessel K, Frey C, Jourdan C, Keil U, et al. Aging sensitizes toward ROS formation and lipid peroxidation in PS1M146L transgenic mice. Free Radic Biol Med 2006;40:850–62.
105. Scripnikov A, Khomenko A, Napryeyenko O; GINDEM-NP Study Group. Effects of Ginkgo biloba extract EGb761 on neuropsychiatric symptoms of dementia: findings from a randomised controlled trial. Wien Med Wochenschr 2007;157:295–300.
106. Shi C, Liu J, Wu F, Yew DT. Ginkgo biloba extract in Alzheimer’s disease: from action mechanisms to medical practice. Int J Mol Sci 2010;11:107–23.
107. Snitz BE, O’Meara ES, Carlson MC, Arnold AM, et al.; Ginkgo Evaluation of Memory (GEM) Study Investigators. Ginkgo biloba for preventing cognitive decline in older adults: a randomized trial. JAMA 2009;302:2663–70.
108. Sokolova L, Hoerr R, Mishchenko T. Treatment of vertigo: A randomized, double-blind trial comparing efficacy and safety of Ginkgo biloba extract EGb761 and betahistine. Int J Otolaryngol 2014;2014:682439.
109. Solomon PR, Adams F, Silver A, Zimmer J, et al. Ginkgo for memory enhancement: a randomized controlled trial. JAMA 2002;288:835–40.
110. Sperling RA, Aisen PS, Beckett LA, Bennett DA, et al. Toward defining the preclinical stages of Alzheimer’s disease: recommendations from the National Institute on Aging-Alzheimer’s Association workgroups on diagnostic guidelines for Alzheimer’s disease. Alzheimers Dement 2011;7: 280–92.
111. Steinberg M, Shao H, Zandi P, Lyketsos CG, et al.; Cache County Investigators. Point and 5-year period prevalence of neuropsychiatric symptoms in dementia: the Cache County Study. Int J Geriatr Psychiatry 2008;23:170–7.
112. Stockburger C, Gold VA, Pallas T, Kolesova N, et al. A cell model for the initial phase of sporadic Alzheimer’s disease. J Alzheimers Dis 2014;42:395–411.
113. Stoll S, Scheuer K, Pohl O, Müller WE. Ginkgo biloba extract (EGb761) independently improves changes in passive avoidance learning and brain membrane fluidity in the aging mouse. Pharmacopsychiatry 1996;29:144–9.
114. Swerdlow RH, Burns JM, Khan SM. The Alzheimer’s disease mitochondrial cascade hypothesis: progress and perspectives. Biochim Biophys Acta 2014;1842:1219–31.
115. Takuma K, Hoshina Y, Arai S, Himeno Y, et al. Ginkgo biloba extract EGb761 attenuates hippocampal neuronal loss and cognitive dysfunction resulting from chronic restraint stress in ovariectomized rats. Neuroscience 2007;149:256–62.
116. Tan MS, Yu JT, Tan CC, Wang HF, et al. Efficacy and adverse effects of ginkgo biloba for cognitive impairment and dementia: a systematic review and meta-analysis. J Alzheimers Dis 2015;43:589–603.
117. Tchantchou F, Xu Y, Wu Y, Christen Y, et al. EGb761 enhances adult hippocampal neurogenesis and phosphorylation of CREB in transgenic mouse model of Alzheimer’s disease. FASEB J 2007;21:2400–8.
118. Tchantchou F, Lacor PN, Cao Z, Lao L, et al. Stimulation of neurogenesis and synaptogenesis by bilobalide and quercetin via common final pathway in hippocampal neurons. J Alzheimers Dis 2009;18:787–98.
119. Ude C, Paulke A, Schubert-Zsilavecz M, Wurglics M. [Chemistry, pharmacokinetics and metabolism of ginkgo extract]. Pharm Unserer Zeit 2009;38:418–23.
120. Vellas B, Coley N, Ousset PJ, Berrut G, et al.; GuidAge Study Group. Long-term use of standardised Ginkgo biloba extract for the prevention of Alzheimer’s disease (GuidAge): a randomised placebo-controlled trial. Lancet Neurol 2012;11:851–9.
121. Vitolo O, Gong B, Cao Z, Ishii H, et al. Protection against beta-amyloid induced abnormal synaptic function and cell death by Ginkgolide J. Neurobiol Aging 2009;30:257–65.
122. von Gunten A, Schlaefke S, Überla K. Efficacy of Ginkgo biloba extract EGb761® in dementia with behavioural and psychological symptoms: A systematic review. World J Biol Psychiatry 2015;(Aug 27):1–12.
123. Walesiuk A, Trofimiuk E, Braszko JJ. Gingko biloba extract diminishes stress-induced memory deficits in rats. Pharmacol Rep 2005;57:176–87.
124. Walesiuk A, Trofimiuk E, Braszko JJ. Ginkgo biloba normalizes stress- and corticosterone-induced impairment of recall in rats. Pharmacol Res 2006;53:123–8.
125. Walesiuk A, Braszko JJ. Preventive action of Ginkgo biloba in stress- and corticosterone-induced impairment of spatial memory in rats. Phytomedicine 2009;16:40–6.
126. Wang BS, Wang H, Song YY, Qi H, et al. Effectiveness of standardized ginkgo biloba extract on cognitive symptoms of dementia with a six-month treatment: a bivariate random effect meta-analysis. Pharmacopsychiatry 2010;43:86–91.
127. Wang Y, Wang L, Wu J, Cai J. The in vivo synaptic plasticity mechanism of EGb 761-induced enhancement of spatial learning and memory in aged rats. Br J Pharmacol 2006;148:147–53.
128. Wang J, Chen W, Wang Y. A ginkgo biloba extract promotes proliferation of endogenous neural stem cells in vascular dementia rats. Neural Regen Res 2013;8:1655–62.
129. Wang ZY, Mo XF, Jiang XH, Rong XF, et al. [Ginkgolide B promotes axonal growth of retina ganglion cells by anti-apoptosis in vitro]. Sheng Li Xue Bao 2012;64:417–24. [Article in Chinese]
130. Weinmann S, Roll S, Schwarzbach C, Vauth C, et al. Effects of Ginkgo biloba in dementia: systematic review and meta-analysis. BMC Geriatr 2010;10:14.
131. Williams B, Watanabe CM, Schultz PG, Rimbach G, et al. Age-related effects of Ginkgo biloba extract on synaptic plasticity and excitability. Neurobiol Aging 2004;25:955–62.
132. Williamson JD, Vellas B, Furberg C, Nahin R, et al. Comparison of the design differences between the Ginkgo Evaluation of Memory study and the GuidAge study. J Nutr Health Aging 2008;12:73S–9S.
133. Winter E. Effects of an extract of Ginkgo biloba on learning and memory in mice. Pharmacol Biochem Behav 1991;38: 109–14.
134. Winter JC. The effects of an extract of Ginkgo biloba, EGb761, on cognitive behavior and longevity in the rat. Physiol Behav 1998;63:425–33.
135. Wirth S, Stemmelin J, Will B, Christen YVES, et al. Facilitative effects of EGb761 on olfactory recognition in young and aged rats. Pharmacol Biochem Behav 2000;65:321–6.
136. Xu Y, Cui C, Pang C, Christen Y, et al. Restoration of impaired phosphorylation of cyclic AMP response element-binding protein (CREB) by EGb761 and its constituents in Abeta-expressing neuroblastoma cells. Eur J Neurosci 2007;26:2931–9.
137. Yancheva S, Ihl R, Nikolova G, Panayotov P, et al.; GINDON Study Group. Ginkgo biloba extract EGb761®, donepezil or both combined in the treatment of Alzheimer’s disease with neuropsychiatric features: a randomised, double-blind, exploratory trial. Aging Ment Health 2009;13:183–90.
138. Yoo DY, Nam Y, Kim W, Yoo KY, et al. Effects of Ginkgo biloba extract on promotion of neurogenesis in the hippocampal dentate gyrus in C57BL/6 mice. J Vet Med Sci 2011;73:71–6. Epub 2010 Aug 30.
139. Yoshitake T, Yoshitake S, Kehr J. The Ginkgo biloba extract EGb761® and its main constituent flavonoids and ginkgolides increase extracellular dopamine levels in the rat prefrontal cortex. Br J Pharmacol 2010;159:659–68.
Dieser Beitrag bezieht sich auf die Vorträge anlässlich des Symposiums „50 Jahre Tebonin – Blick nach Vorn“ organisiert von der Hirnliga e.V. und der Dr. Willmar Schwabe GmbH & Co. an der Universität Frankfurt im Januar 2015.
Univ.-Prof. Dr. Walter E. Müller, Höhenstraße 49A, 67550 Worms und Pharmakologisches Institut für Naturwissenschaftler, Goethe-Universität, Biozentrum, Frankfurt/M., E-Mail: w.e.mueller@em.uni-frankfurt.de
Prof. Dr. Anne Eckert, Neurobiologisches Forschungslabor, Psychiatrische Universitätsklinik, Basel
Prof. Dr. Gunter P. Eckert, Justus-Liebig- Universität, Institut für Ernährungswissenschaften (IfE), Wilhelmstraße 20, 35392 Gießen.
Prof. Dr. Heidrun Fink, Institut für Pharmakologie und Toxikologie, Freie Universität Berlin, Berlin
Prof. Dr. Kristina Friedland, Institut für Molekulare und Klinische Pharmazie, Universität Erlangen, Erlangen
Dr.med. Robert Hörr, Klinische Forschung, Geriatrie/ZNS, Dr. Willmar Schwabe GmbH & Co. KG, Karlsruhe
Prof. Dr.med. Ralf Ihl, Alexianer Krankenhaus, Krefeld
O. Univ.-Prof. Dr. Siegfried Kasper, Universitätsklinik für Psychiatrie – AKH, Wien
Prof. Dr.med. Hans-Jürgen Möller, Psychiatrische Klinik und Poliklinik, LMU München
The Ginkgo special extract EGb761® (Tebonin®). A preclinical and clinical update in respect to the different clinical and neurobiological concepts of Alzheimer‘s disease
The Ginkgo biloba leaves’ special extract EGb761® was introduced into therapy in Germany 50 years ago under the brand name Tebonin®. Even from the beginning it was used for age-associated cognitive impairment, originally under the diagnosis organic brain syndrome and later fort the diagnosis of dementia either of Alzheimer or vascular type. However, because of methodological problems of several of the older clinical studies, there was always a controversy about its use in the treatment of dementia. The sceptical appraisal was also associated with the mechanism of action of EGb761®, an improvement of mitochondrial function leading to enhanced neuronal function and improved neuronal plasticity, since these effects were not specific for Alzheimer pathology but also relevant for age-associated memory impairment. However, due to our actual knowledge about mitochondrial dysfunction as a major cause of the whole continuum of age-associated memory disorders including Alzheimer’s disease, it became clear that a drug which improves mitochondrial function should be effective over the whole spectrum, from mild cognitive disturbances up to Alzheimer dementia. Accordingly, a positive assessment of EGb761® for the treatment of dementia has been included into the guidelines for the treatment of Alzheimer and other dementias of the World Societies of Biological Psychiatry and in the German „S3-Leitlinie Demenz“.
Key words: Ginkgo, EGb761®, mitochondrial dysfunction, treatment of dementia
Psychopharmakotherapie 2016; 23(03)